

A Novel Interaction between Complement Inhibitor C4b-binding Protein and Plasminogen That Enhances Plasminogen Activation
- PMID: 26067271
- PMCID: PMC4513095
- DOI: 10.1074/jbc.M114.619494
A Novel Interaction between Complement Inhibitor C4b-binding Protein and Plasminogen That Enhances Plasminogen Activation
Abstract
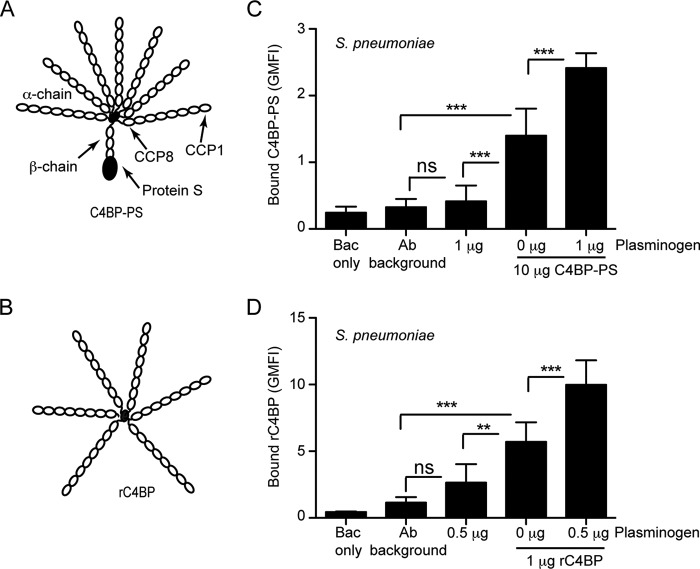
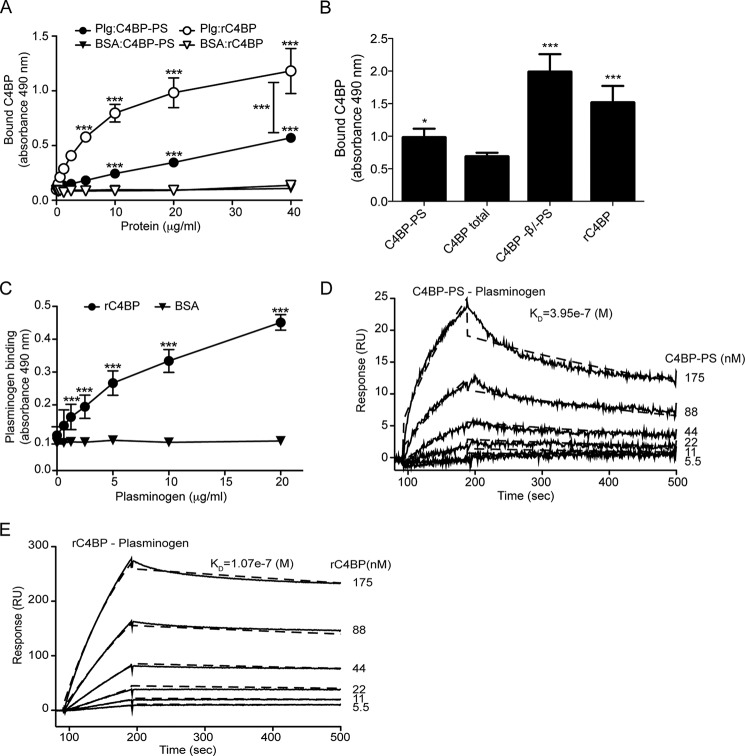
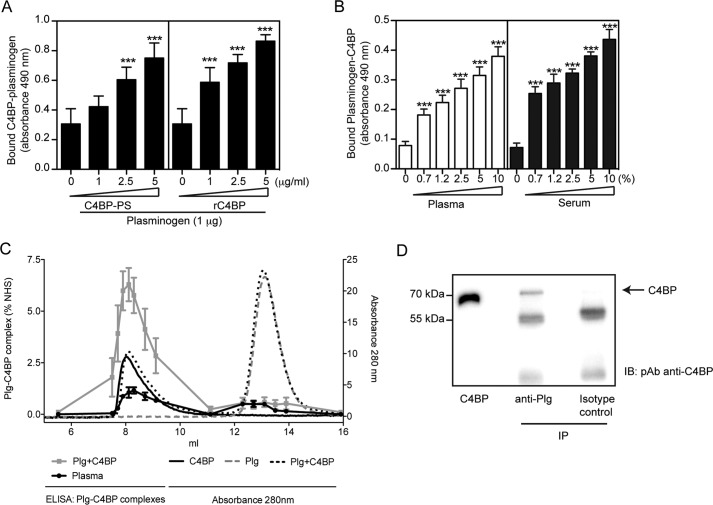
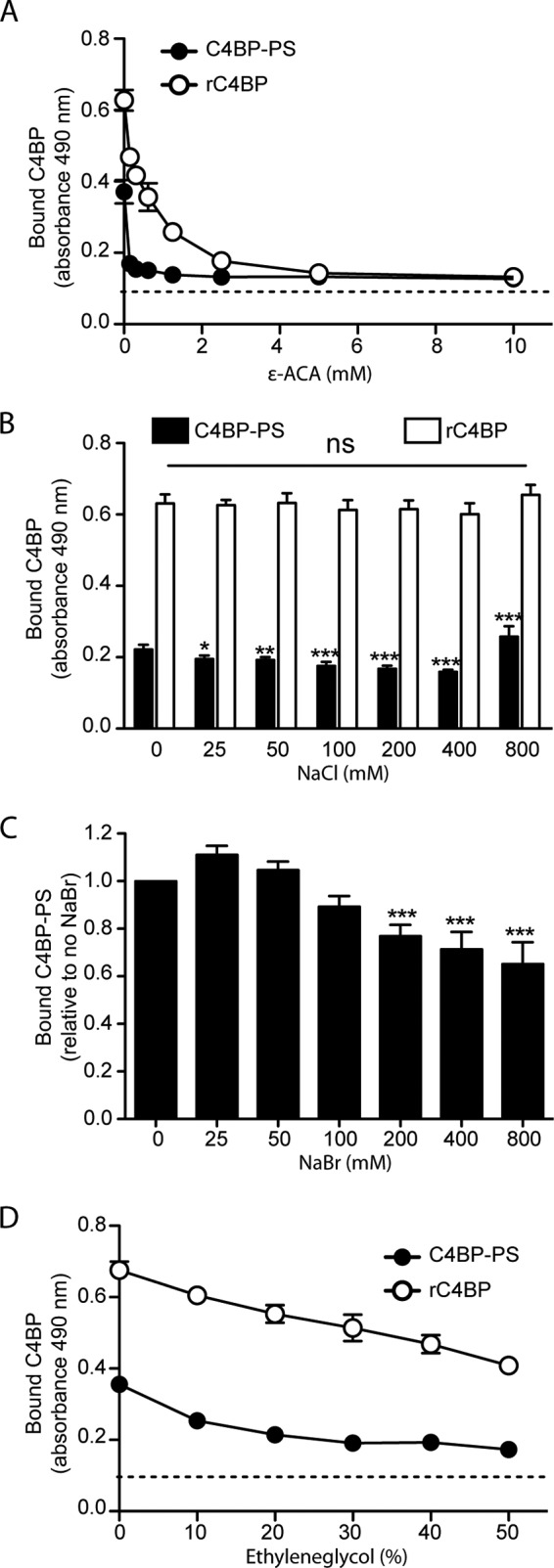
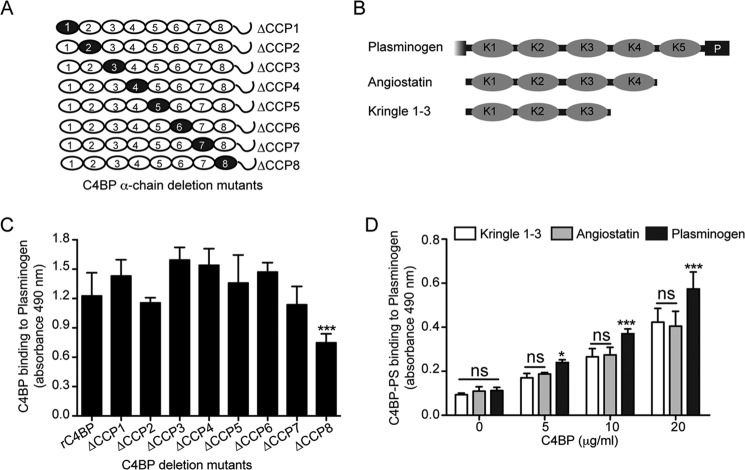
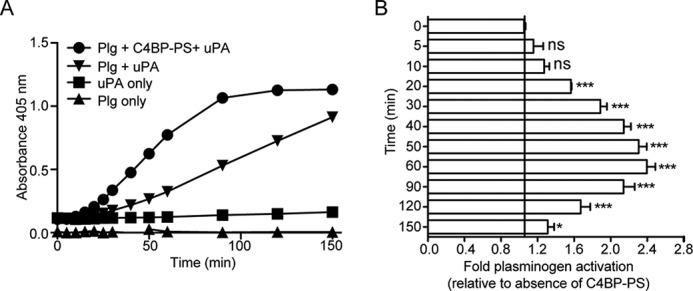
The complement, coagulation, and fibrinolytic systems are crucial for the maintenance of tissue homeostasis. To date numerous interactions and cross-talks have been identified between these cascades. In line with this, here we propose a novel, hitherto unknown interaction between the complement inhibitor C4b-binding protein (C4BP) and plasminogen of the fibrinolytic pathway. Binding of C4BP to Streptococcus pneumoniae is a known virulence mechanism of this pathogen and it was increased in the presence of plasminogen. Interestingly, the acute phase variant of C4BP lacking the β-chain and protein S binds plasminogen much stronger than the main isoform containing the β-chain and protein S. Indeed, the complement control protein (CCP) 8 domain of C4BP, which would otherwise be sterically hindered by the β-chain, primarily mediates this interaction. Moreover, the lysine-binding sites in plasminogen kringle domains facilitate the C4BP-plasminogen interaction. Furthermore, C4BP readily forms complexes with plasminogen in fluid phase and such complexes are present in human serum and plasma. Importantly, whereas the presence of plasminogen did not affect the factor I cofactor activity of C4BP, the activation of plasminogen by urokinase-type plasminogen activator to active plasmin was significantly augmented in the presence of C4BP. Taken together, our data demonstrate a novel interaction between two proteins of the complement and fibrinolytic system. Most complexes might be formed during the acute phase of inflammation and have an effect on the homeostasis at the site of injury or acute inflammation.
Keywords: C4b-binding protein; complement; plasmin; plasminogen; protein-protein interaction; surface plasmon resonance (SPR).
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Holers V. M. (2014) Complement and its receptors: new insights into human disease. Annu. Rev. Immunol. 32, 433–459 - PubMed
-
- Sjöberg A. P., Trouw L. A., Blom A. M. (2009) Complement activation and inhibition: a delicate balance. Trends Immunol. 30, 83–90 - PubMed
-
- Zipfel P. F., Skerka C. (2009) Complement regulators and inhibitory proteins. Nat. Rev. Immunol. 9, 729–740 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
