

PHYSIOLOGY. Regulation of breathing by CO₂ requires the proton-activated receptor GPR4 in retrotrapezoid nucleus neurons
- PMID: 26068853
- PMCID: PMC5171229
- DOI: 10.1126/science.aaa0922
PHYSIOLOGY. Regulation of breathing by CO₂ requires the proton-activated receptor GPR4 in retrotrapezoid nucleus neurons
Abstract
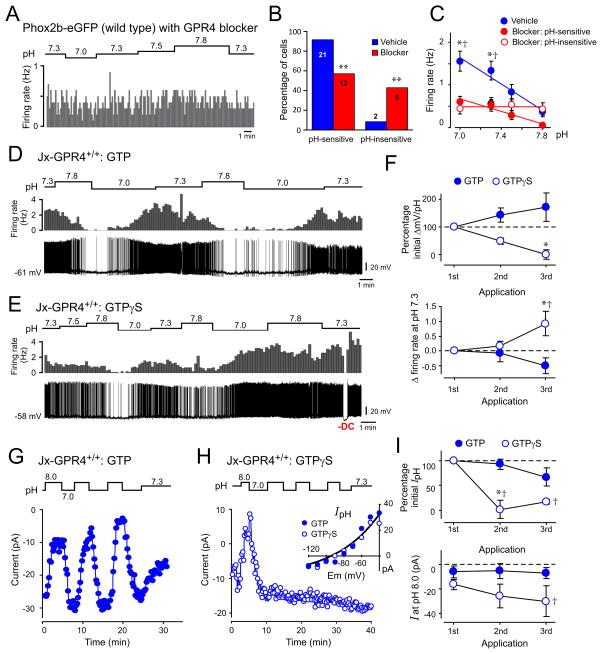
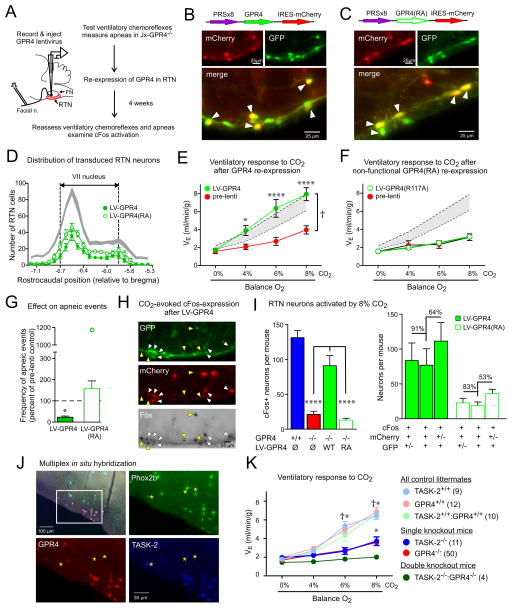
Blood gas and tissue pH regulation depend on the ability of the brain to sense CO2 and/or H(+) and alter breathing appropriately, a homeostatic process called central respiratory chemosensitivity. We show that selective expression of the proton-activated receptor GPR4 in chemosensory neurons of the mouse retrotrapezoid nucleus (RTN) is required for CO2-stimulated breathing. Genetic deletion of GPR4 disrupted acidosis-dependent activation of RTN neurons, increased apnea frequency, and blunted ventilatory responses to CO2. Reintroduction of GPR4 into RTN neurons restored CO2-dependent RTN neuronal activation and rescued the ventilatory phenotype. Additional elimination of TASK-2 (K(2P)5), a pH-sensitive K(+) channel expressed in RTN neurons, essentially abolished the ventilatory response to CO2. The data identify GPR4 and TASK-2 as distinct, parallel, and essential central mediators of respiratory chemosensitivity.
Copyright © 2015, American Association for the Advancement of Science.
Figures


References
-
- Mulkey DK, et al. Respiratory control by ventral surface chemoreceptor neurons in rats. Nat Neurosci. 2004;7 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
