

IL-17A promotes protective IgA responses and expression of other potential effectors against the lumen-dwelling enteric parasite Giardia
- PMID: 26071205
- PMCID: PMC4547885
- DOI: 10.1016/j.exppara.2015.06.003
IL-17A promotes protective IgA responses and expression of other potential effectors against the lumen-dwelling enteric parasite Giardia
Abstract
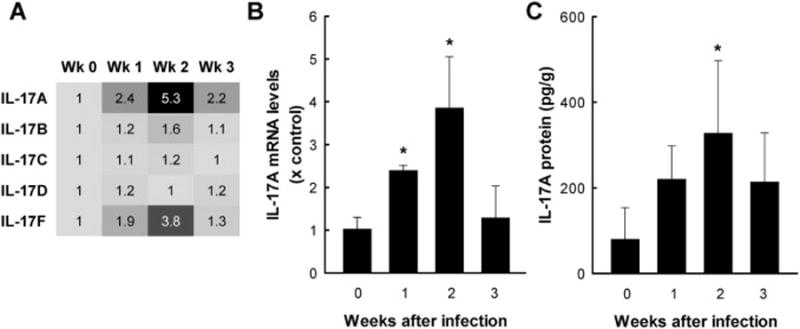
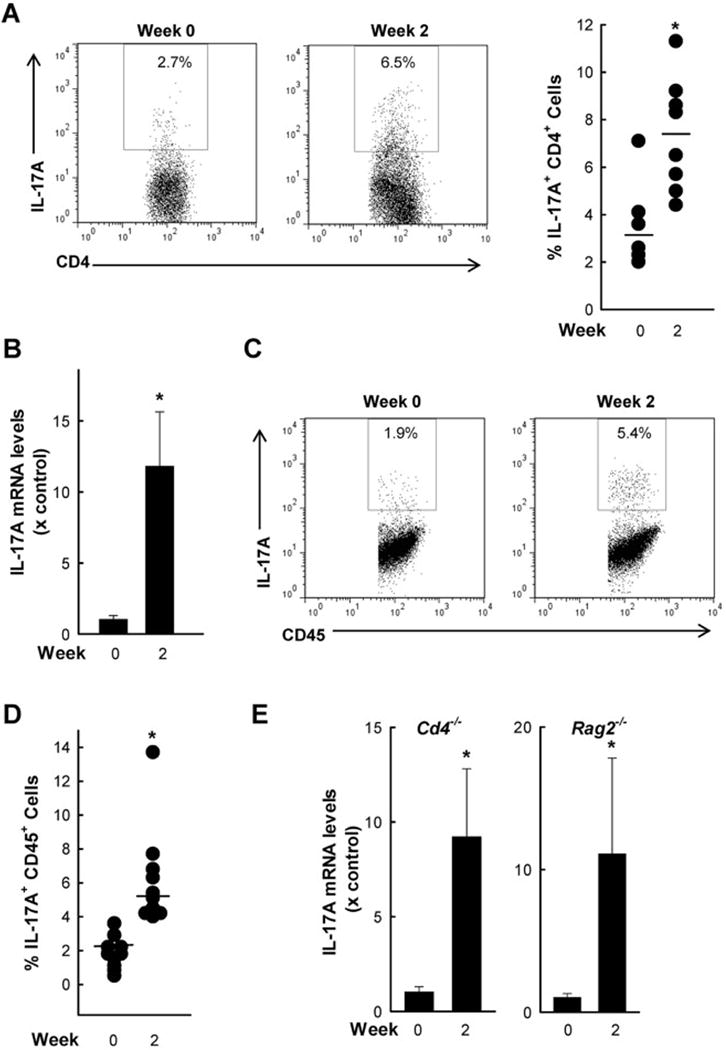
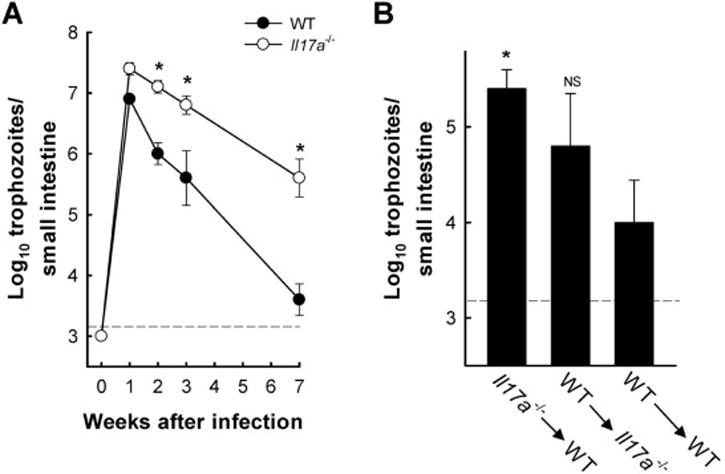
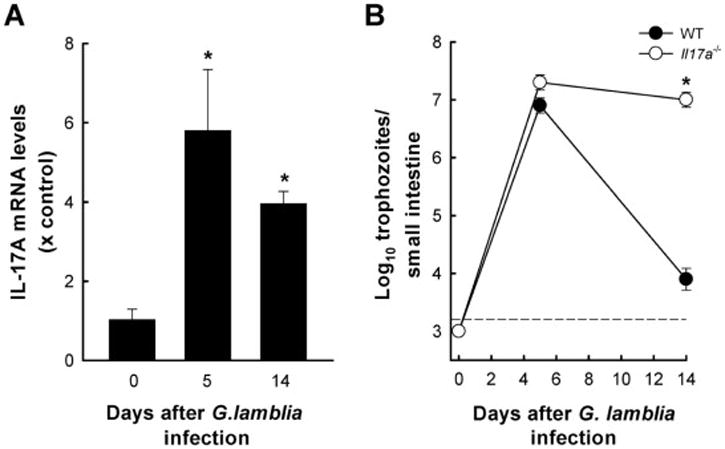
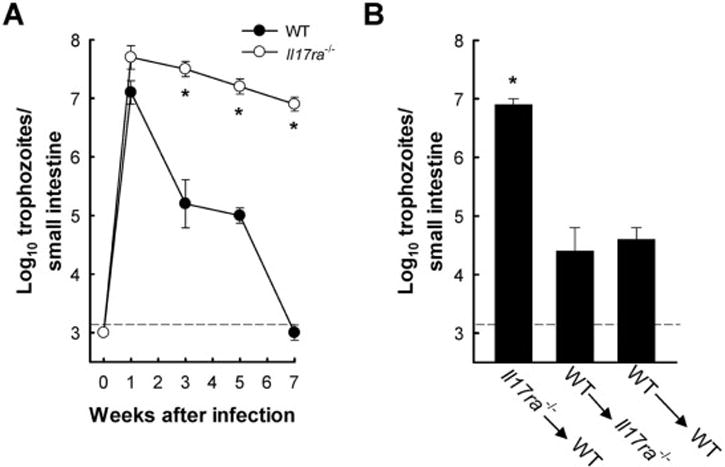
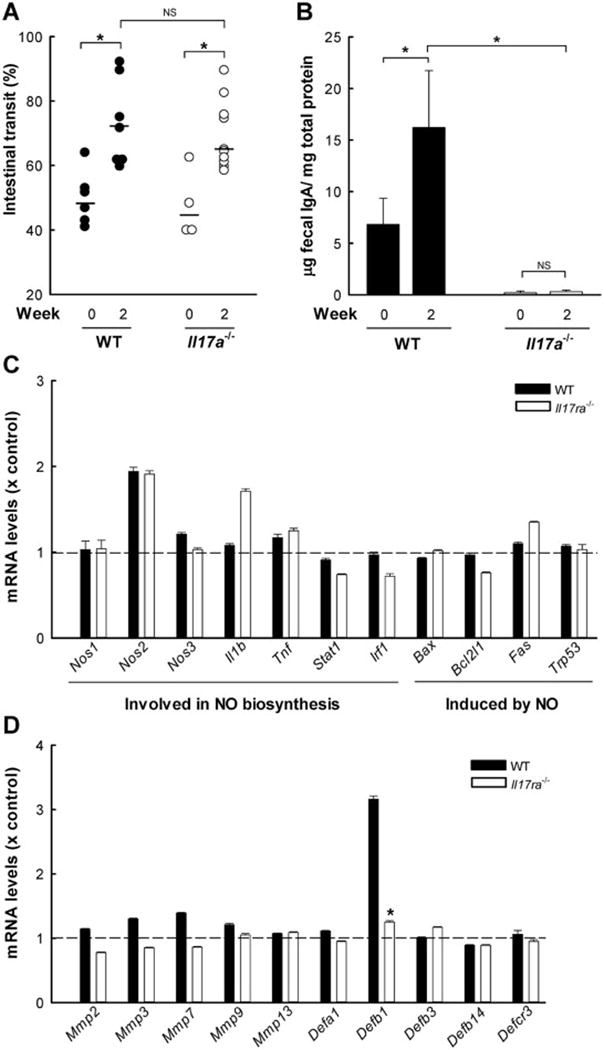
Giardia lamblia is a leading protozoan cause of diarrheal disease worldwide. It colonizes the lumen and epithelial surface of the small intestine, but does not invade the mucosa. Acute infection causes only minimal mucosal inflammation. Effective immune defenses exist, yet their identity and mechanisms remain incompletely understood. Interleukin (IL)-17A has emerged as an important cytokine involved in inflammation and antimicrobial defense against bacterial pathogens at mucosal surfaces. In this study, we demonstrate that IL-17A has a crucial function in host defense against Giardia infection. Using murine infection models with G. muris and G. lamblia, we observed marked and selective induction of intestinal IL-17A with peak expression after 2 weeks. Th17 cells in the lamina propria and innate immune cells in the epithelial compartment of the small intestine were responsible for the IL-17A response. Experiments in gene-targeted mice revealed that the cytokine, and its cognate receptor IL-17RA, were required for eradication of the parasite. The actions of the cytokine were mediated by hematopoietic cells, and were required for the transport of IgA into the intestinal lumen, since IL-17A deficiency led to marked reduction of fecal IgA levels, as well as for increased intestinal expression of several other potential effectors, including β-defensin 1 and resistin-like molecule β. In contrast, intestinal hypermotility, another major antigiardial defense mechanism, was not impacted by IL-17A loss. Taken together, these findings demonstrate that IL-17A and IL-17 receptor signaling are essential for intestinal defense against the important lumen-dwelling intestinal parasite Giardia.
Keywords: Cytokines; Mucosal immunity; Protozoan parasites; Small intestine.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
