

ER Stress Sensor XBP1 Controls Anti-tumor Immunity by Disrupting Dendritic Cell Homeostasis
- PMID: 26073941
- PMCID: PMC4580135
- DOI: 10.1016/j.cell.2015.05.025
ER Stress Sensor XBP1 Controls Anti-tumor Immunity by Disrupting Dendritic Cell Homeostasis
Abstract
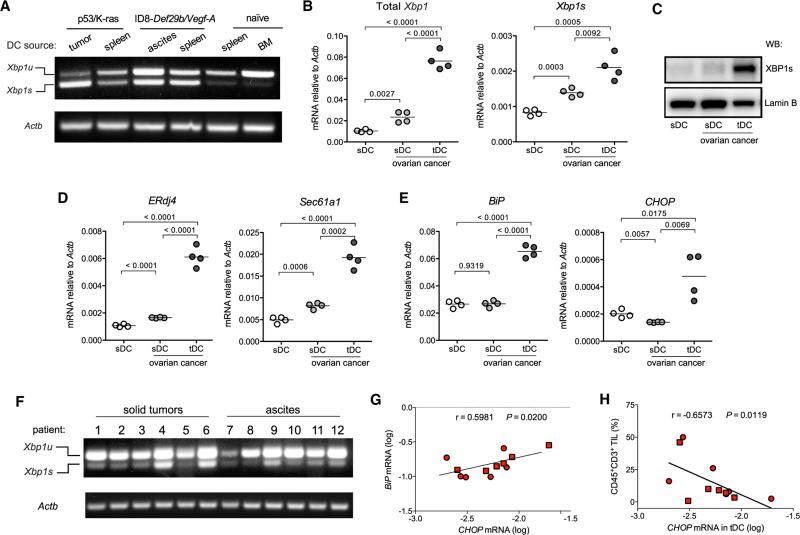
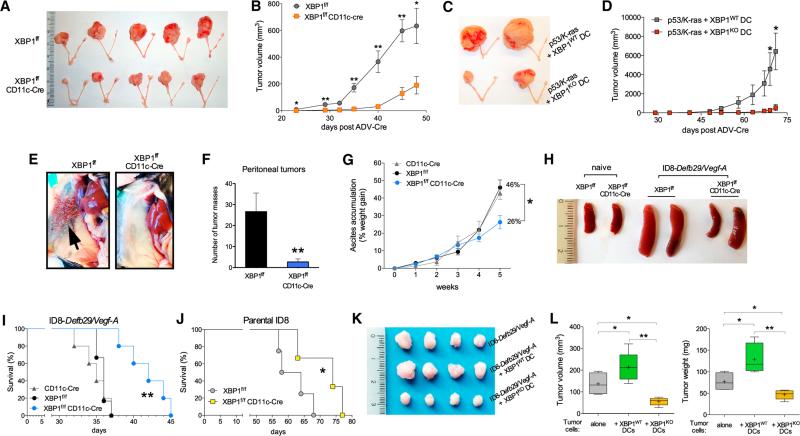
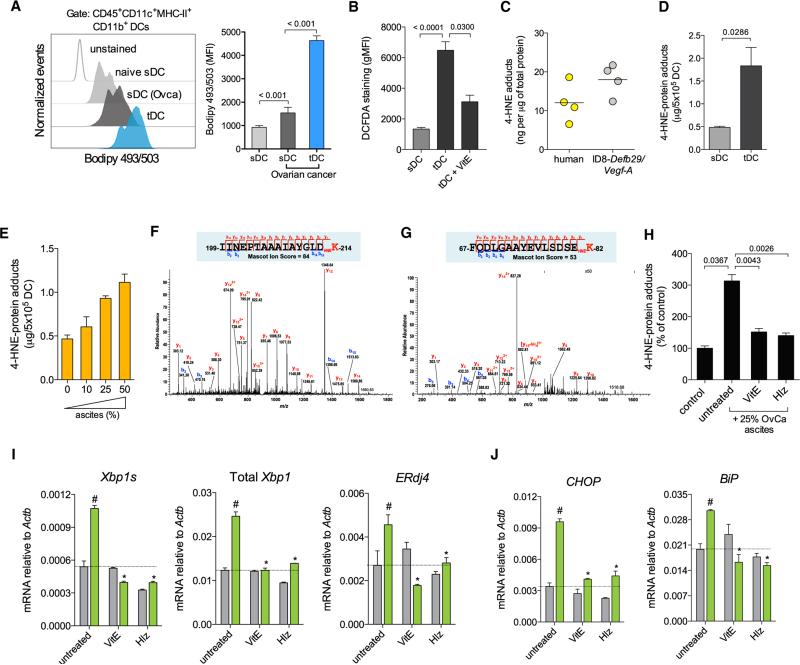
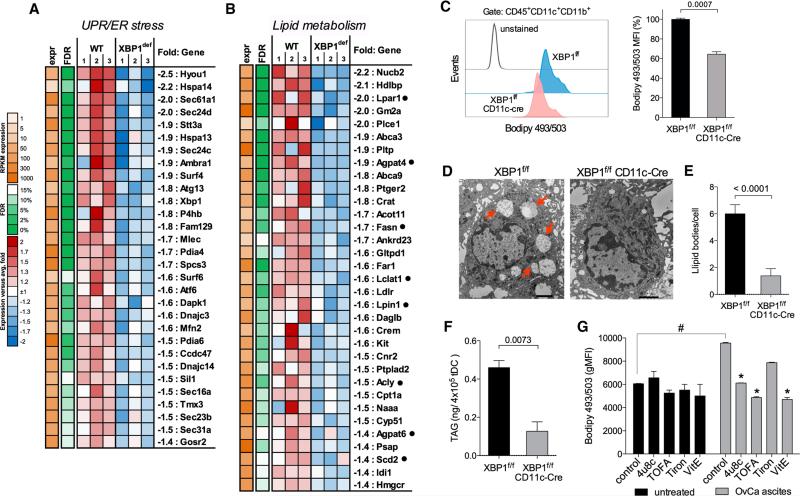
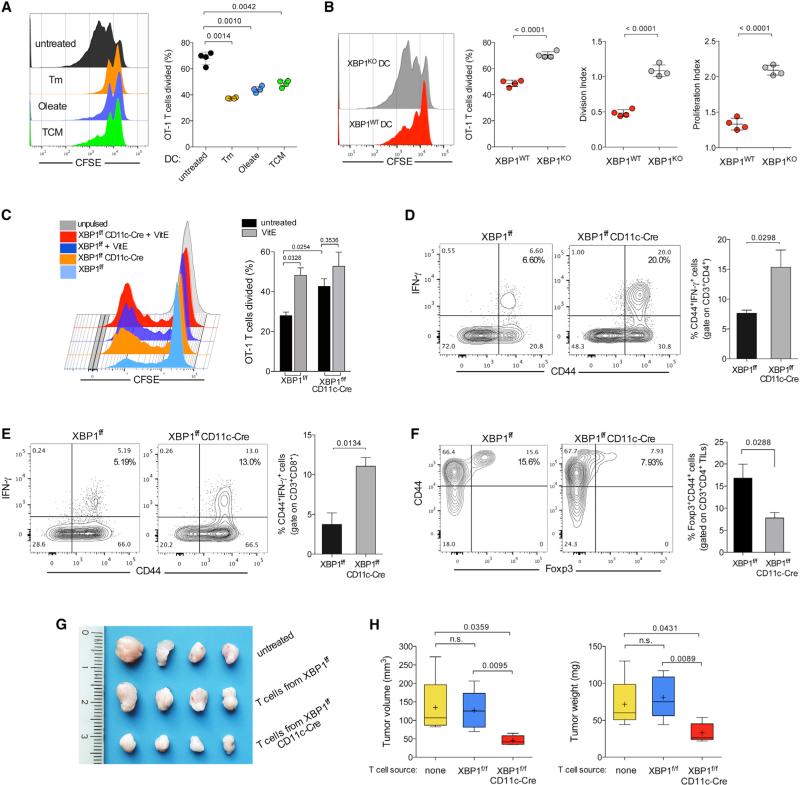
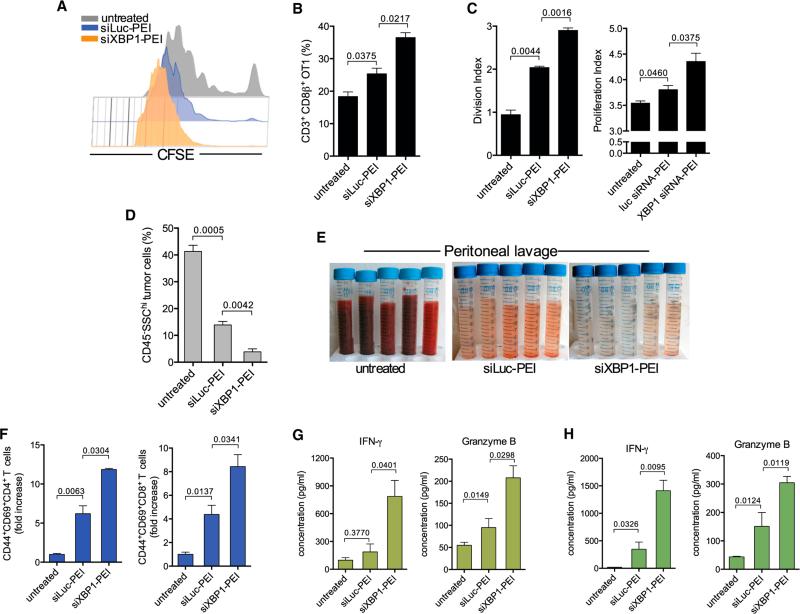
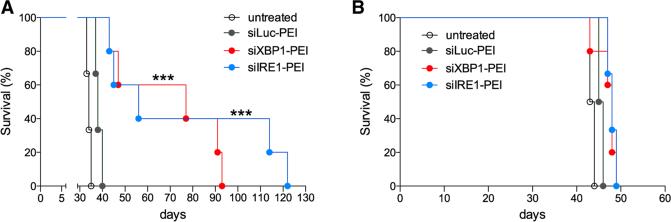
Dendritic cells (DCs) are required to initiate and sustain T cell-dependent anti-cancer immunity. However, tumors often evade immune control by crippling normal DC function. The endoplasmic reticulum (ER) stress response factor XBP1 promotes intrinsic tumor growth directly, but whether it also regulates the host anti-tumor immune response is not known. Here we show that constitutive activation of XBP1 in tumor-associated DCs (tDCs) drives ovarian cancer (OvCa) progression by blunting anti-tumor immunity. XBP1 activation, fueled by lipid peroxidation byproducts, induced a triglyceride biosynthetic program in tDCs leading to abnormal lipid accumulation and subsequent inhibition of tDC capacity to support anti-tumor T cells. Accordingly, DC-specific XBP1 deletion or selective nanoparticle-mediated XBP1 silencing in tDCs restored their immunostimulatory activity in situ and extended survival by evoking protective type 1 anti-tumor responses. Targeting the ER stress response should concomitantly inhibit tumor growth and enhance anti-cancer immunity, thus offering a unique approach to cancer immunotherapy.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
ER Stress in Dendritic Cells Promotes Cancer.Cell. 2015 Jun 18;161(7):1492-3. doi: 10.1016/j.cell.2015.06.006. Cell. 2015. PMID: 26091029
-
Tumour immunology: Stressed DCs can't handle T cells.Nat Rev Immunol. 2015 Aug;15(8):465. doi: 10.1038/nri3885. Epub 2015 Jul 3. Nat Rev Immunol. 2015. PMID: 26139351 No abstract available.
-
Cancer: A dendritic-cell brake on antitumour immunity.Nature. 2015 Jul 16;523(7560):294-5. doi: 10.1038/523294a. Nature. 2015. PMID: 26178959 Free PMC article.
-
Dendritic cells are stressed out in tumor.Cell Res. 2015 Sep;25(9):989-90. doi: 10.1038/cr.2015.93. Epub 2015 Jul 31. Cell Res. 2015. PMID: 26227962 Free PMC article.
References
-
- Acosta-Alvear D, Zhou Y, Blais A, Tsikitis M, Lents NH, Arias C, Lennon CJ, Kluger Y, Dynlacht BD. XBP1 controls diverse cell type- and condition-specific transcriptional regulatory networks. Mol. Cell. 2007;27:53–66. - PubMed
-
- Conejo-Garcia JR, Benencia F, Courreges MC, Kang E, Mohamed-Hadley A, Buckanovich RJ, Holtz DO, Jenkins A, Na H, Zhang L, et al. Tumor-infiltrating dendritic cell precursors recruited by a beta-defensin contribute to vasculogenesis under the influence of Vegf-A. Nat. Med. 2004;10:950–958. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
