

Sleep Facilitates Memory by Blocking Dopamine Neuron-Mediated Forgetting
- PMID: 26073942
- PMCID: PMC4671826
- DOI: 10.1016/j.cell.2015.05.027
Sleep Facilitates Memory by Blocking Dopamine Neuron-Mediated Forgetting
Abstract
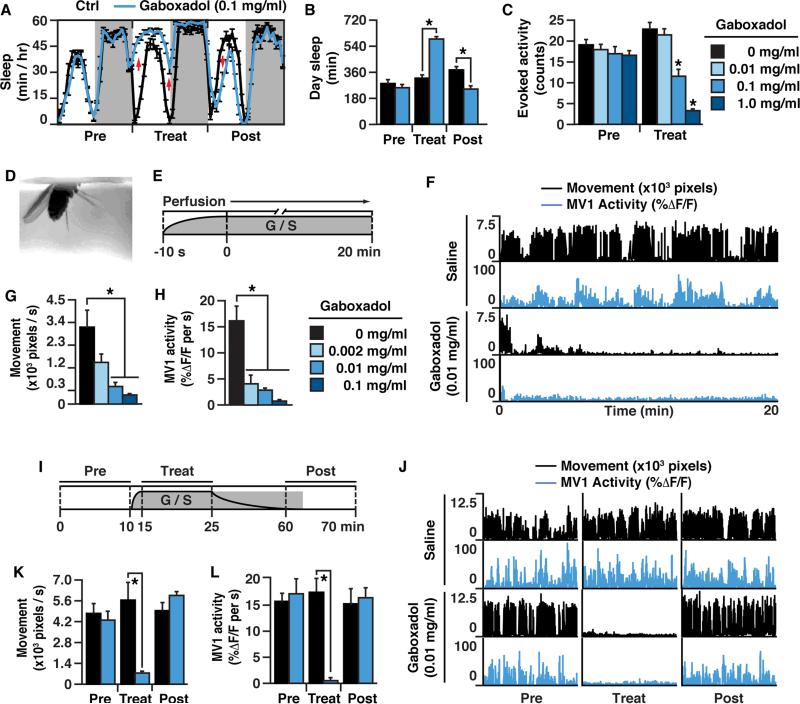
Early studies from psychology suggest that sleep facilitates memory retention by stopping ongoing retroactive interference caused by mental activity or external sensory stimuli. Neuroscience research with animal models, on the other hand, suggests that sleep facilitates retention by enhancing memory consolidation. Recently, in Drosophila, the ongoing activity of specific dopamine neurons was shown to regulate the forgetting of olfactory memories. Here, we show this ongoing dopaminergic activity is modulated with behavioral state, increasing robustly with locomotor activity and decreasing with rest. Increasing sleep-drive, with either the sleep-promoting agent Gaboxadol or by genetic stimulation of the neural circuit for sleep, decreases ongoing dopaminergic activity, while enhancing memory retention. Conversely, increasing arousal stimulates ongoing dopaminergic activity and accelerates dopaminergic-based forgetting. Therefore, forgetting is regulated by the behavioral state modulation of dopaminergic-based plasticity. Our findings integrate psychological and neuroscience research on sleep and forgetting.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Flies Sleep on It, or Fuhgeddaboudit!Cell. 2015 Jun 18;161(7):1498-500. doi: 10.1016/j.cell.2015.06.011. Cell. 2015. PMID: 26091032
-
Sleep: Don't forget.Nat Rev Neurosci. 2015 Aug;16(8):442-3. doi: 10.1038/nrn3990. Epub 2015 Jul 1. Nat Rev Neurosci. 2015. PMID: 26130162 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
