

Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection
- PMID: 26074005
- PMCID: PMC4631312
- DOI: 10.1016/j.neuron.2015.05.038
Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection
Abstract
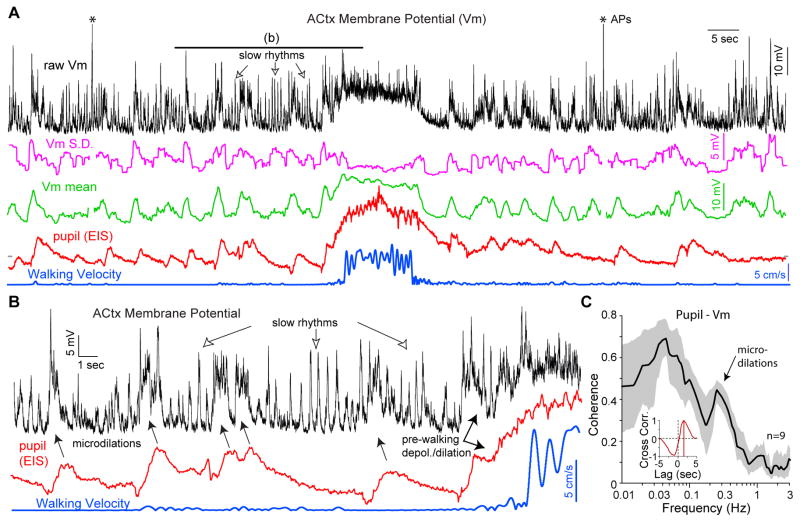
The neural correlates of optimal states for signal detection task performance are largely unknown. One hypothesis holds that optimal states exhibit tonically depolarized cortical neurons with enhanced spiking activity, such as occur during movement. We recorded membrane potentials of auditory cortical neurons in mice trained on a challenging tone-in-noise detection task while assessing arousal with simultaneous pupillometry and hippocampal recordings. Arousal measures accurately predicted multiple modes of membrane potential activity, including rhythmic slow oscillations at low arousal, stable hyperpolarization at intermediate arousal, and depolarization during phasic or tonic periods of hyper-arousal. Walking always occurred during hyper-arousal. Optimal signal detection behavior and sound-evoked responses, at both sub-threshold and spiking levels, occurred at intermediate arousal when pre-decision membrane potentials were stably hyperpolarized. These results reveal a cortical physiological signature of the classically observed inverted-U relationship between task performance and arousal and that optimal detection exhibits enhanced sensory-evoked responses and reduced background synaptic activity.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures







Comment in
-
Behavioral State--Getting "In The Zone".Neuron. 2015 Jul 1;87(1):7-9. doi: 10.1016/j.neuron.2015.06.020. Neuron. 2015. PMID: 26139365 Free PMC article.
References
-
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
