

Beyond reward prediction errors: the role of dopamine in movement kinematics
- PMID: 26074791
- PMCID: PMC4444742
- DOI: 10.3389/fnint.2015.00039
Beyond reward prediction errors: the role of dopamine in movement kinematics
Abstract
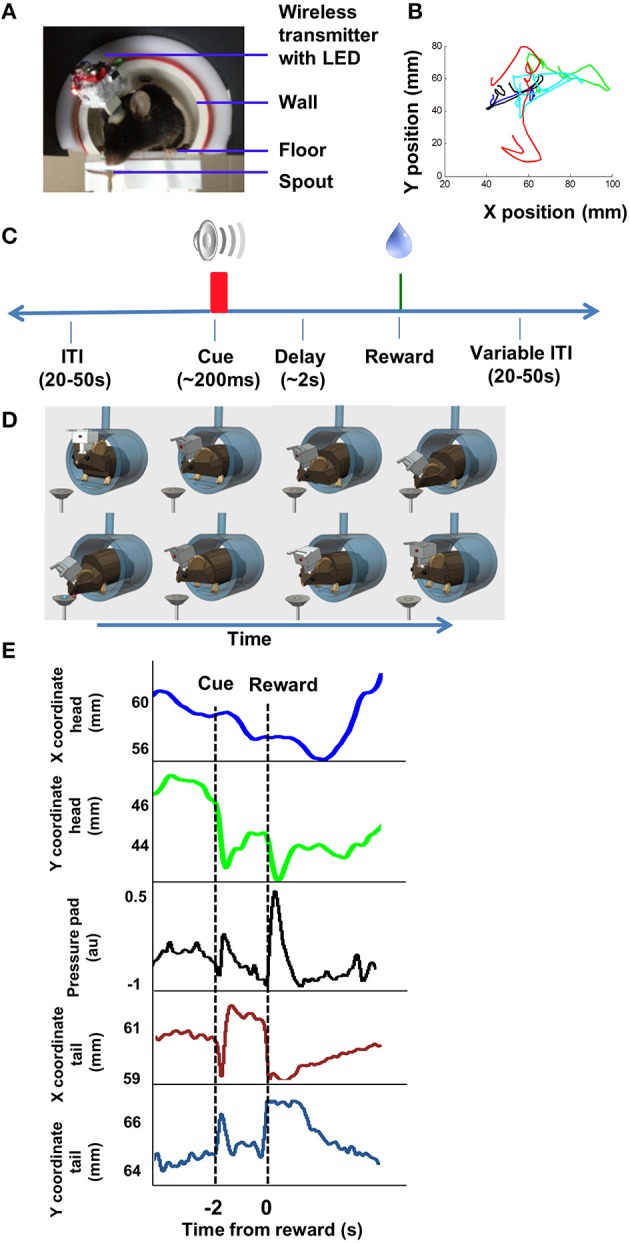
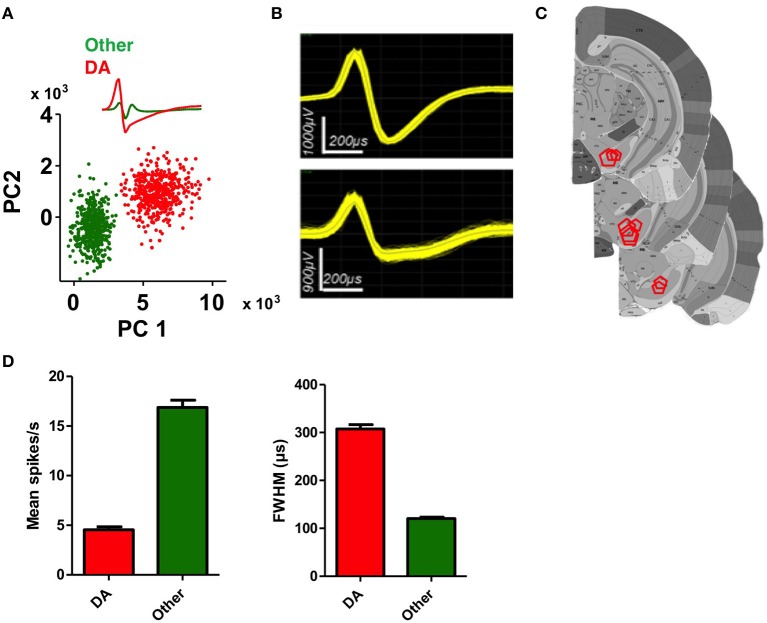
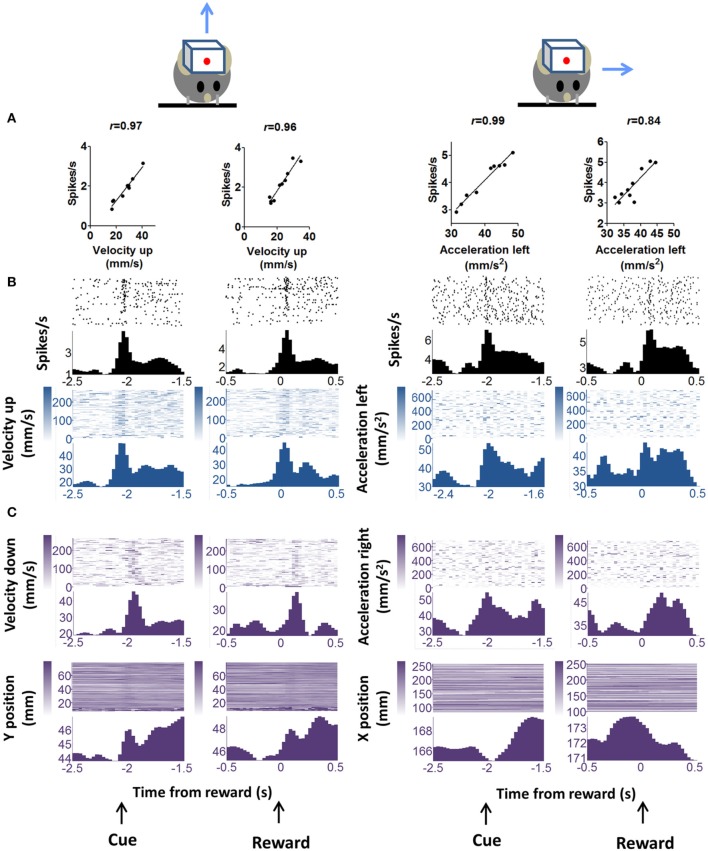
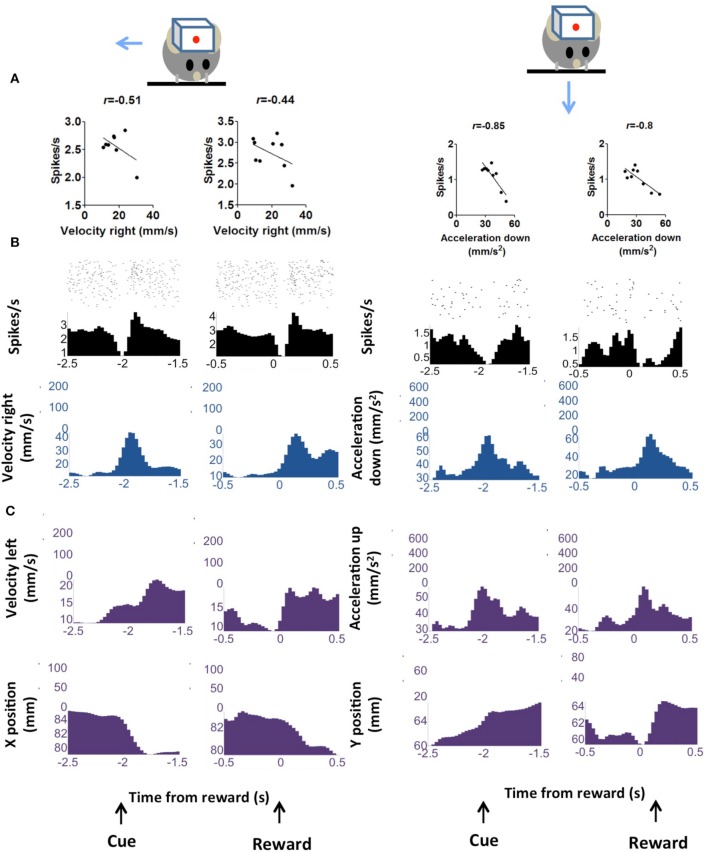
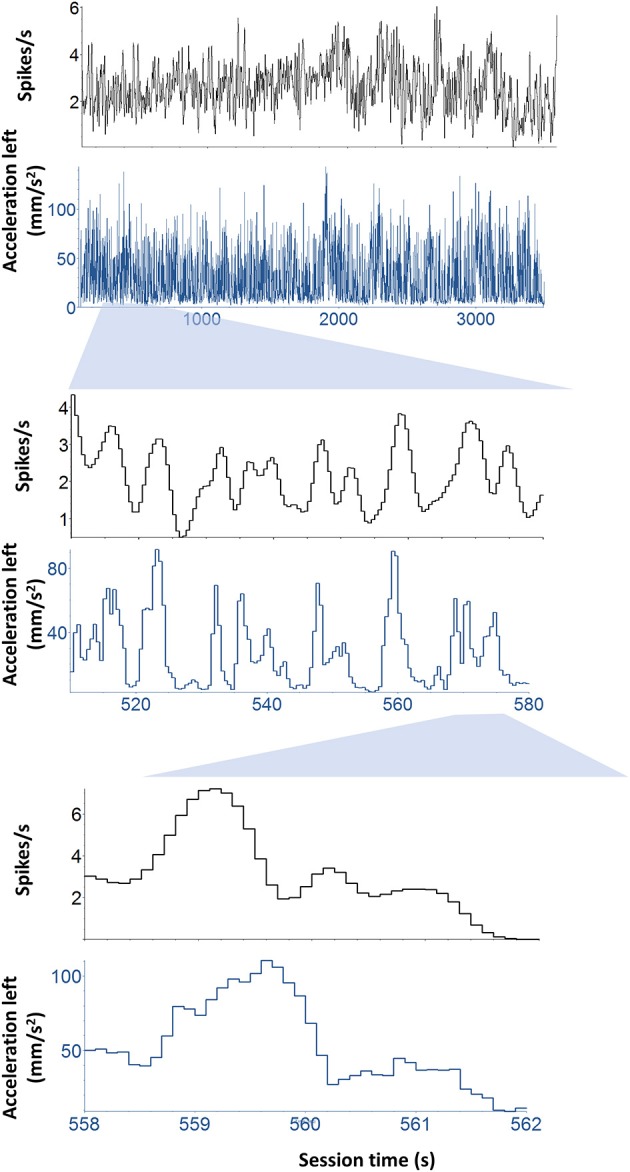
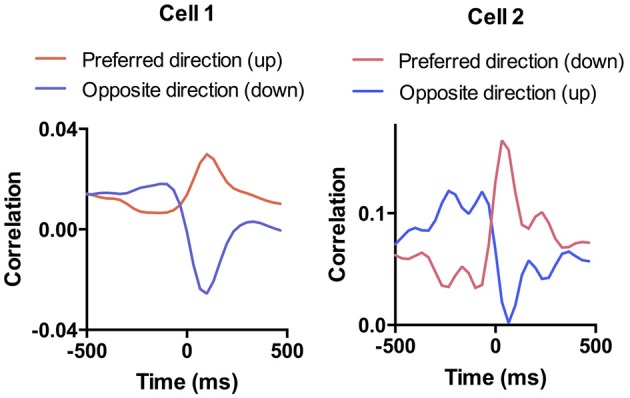
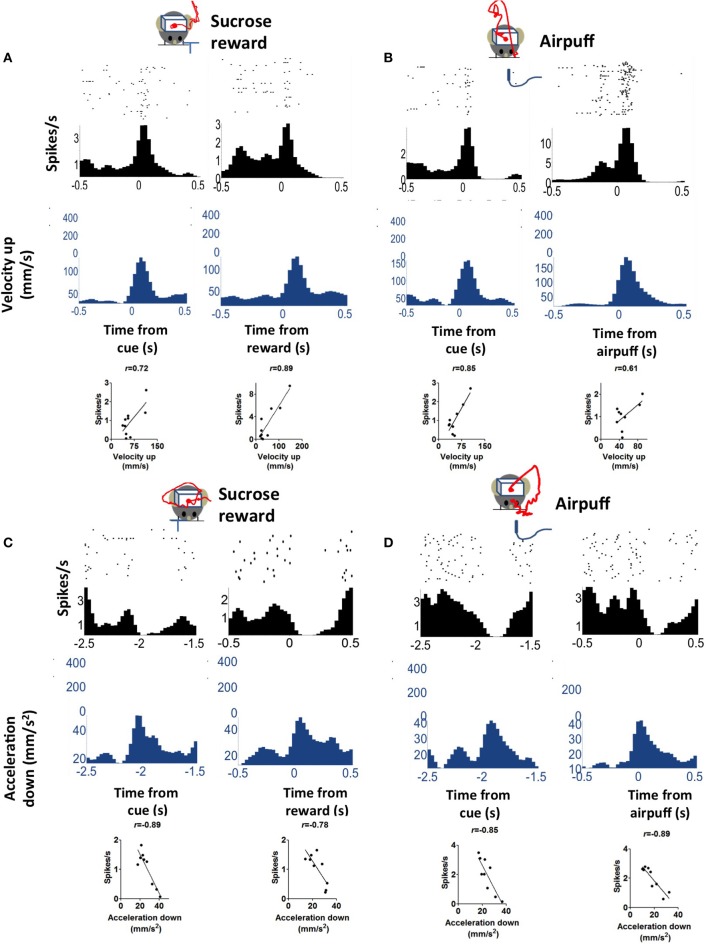
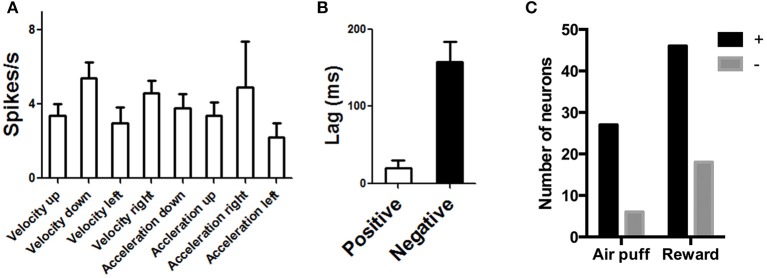
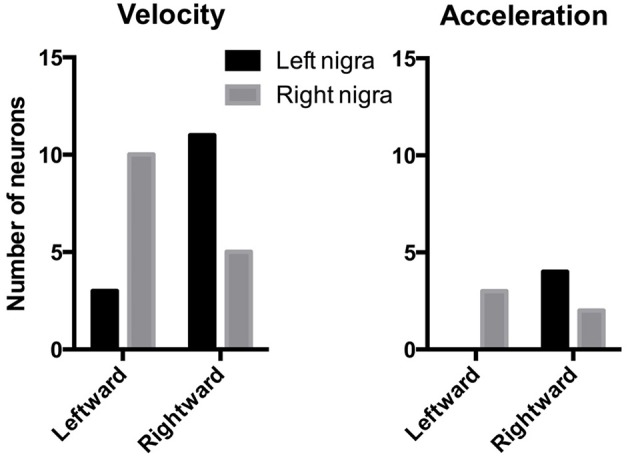
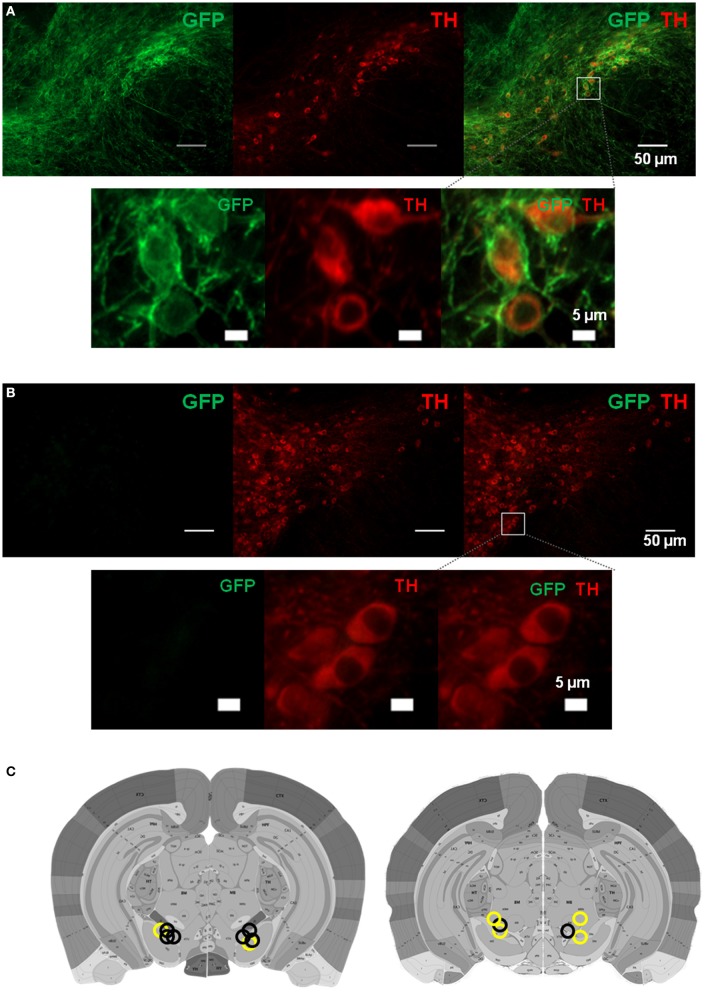
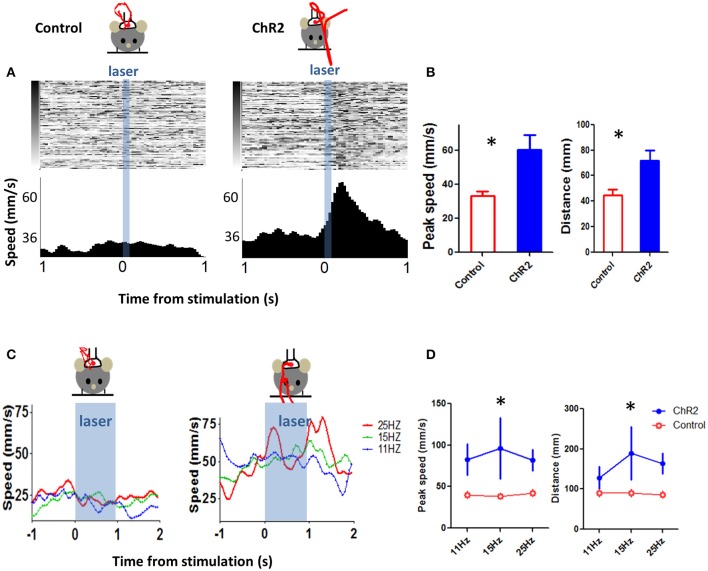
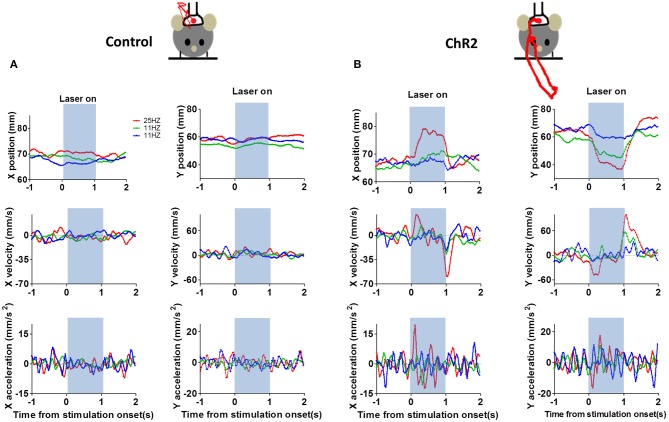
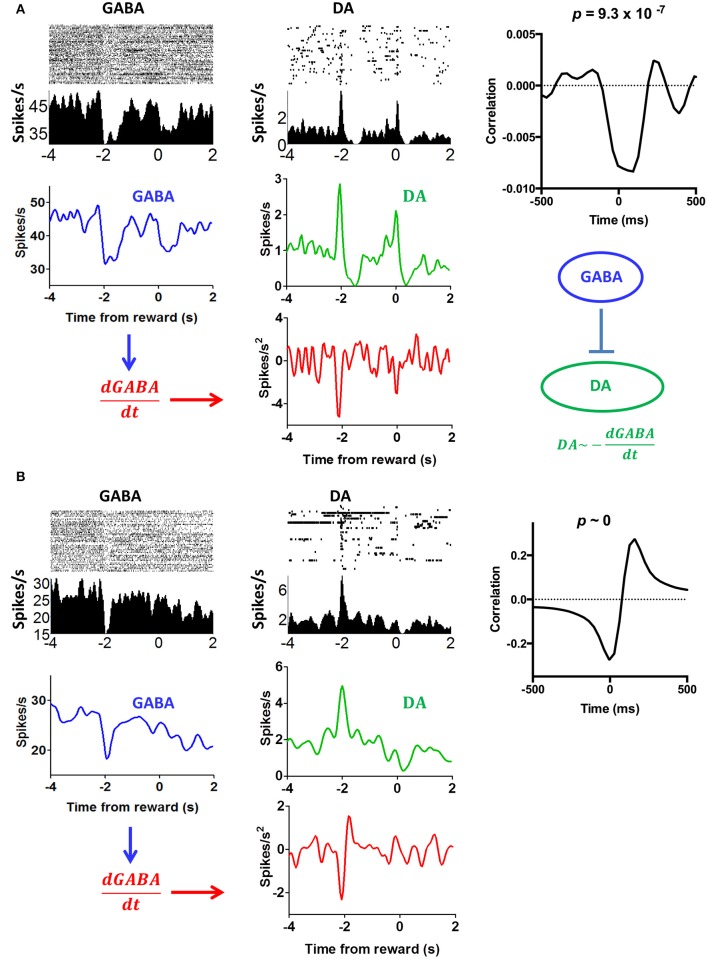
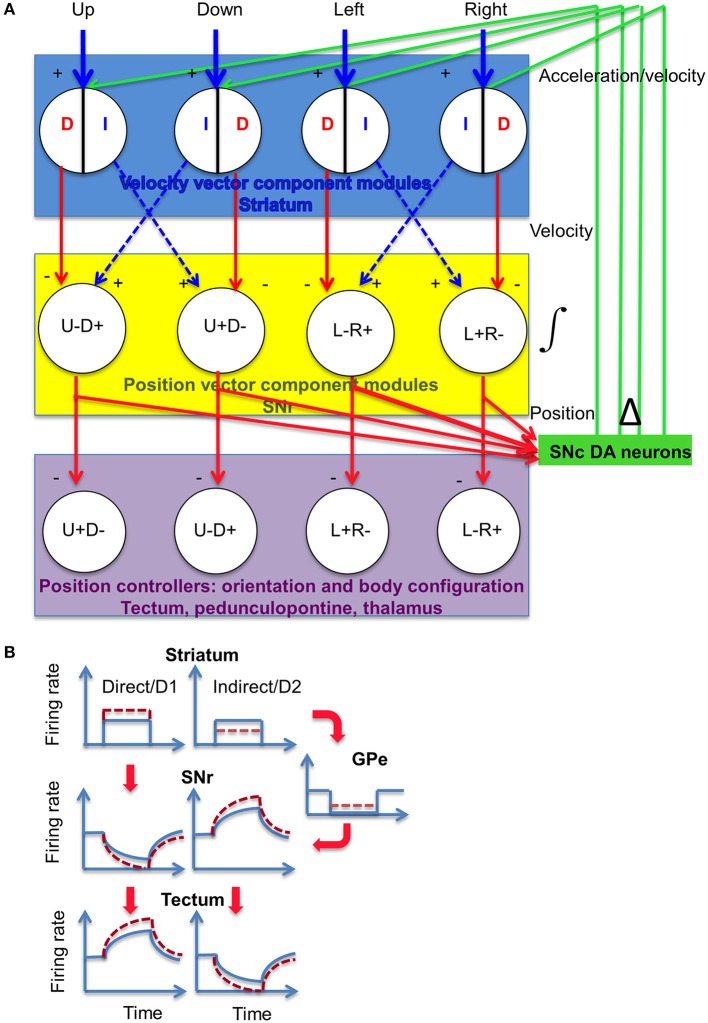
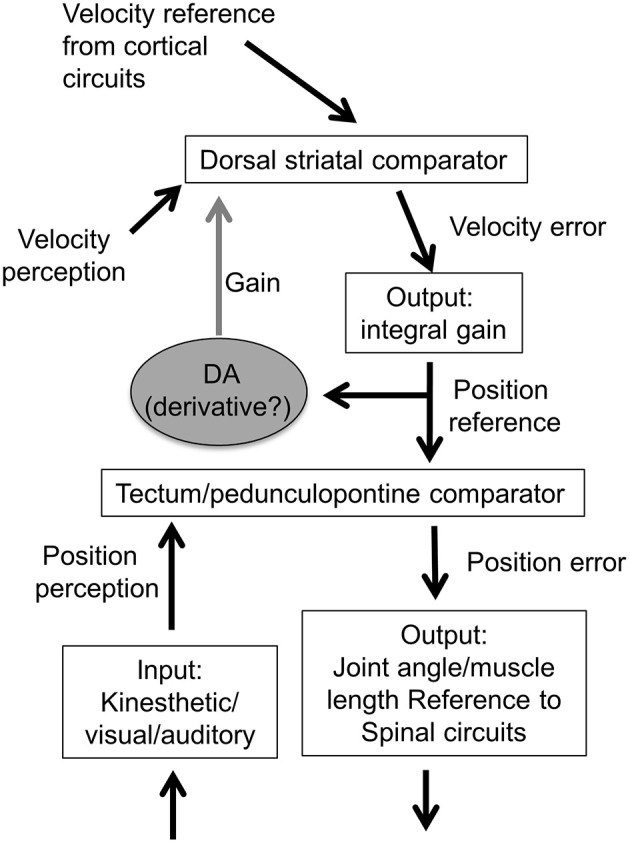
We recorded activity of dopamine (DA) neurons in the substantia nigra pars compacta in unrestrained mice while monitoring their movements with video tracking. Our approach allows an unbiased examination of the continuous relationship between single unit activity and behavior. Although DA neurons show characteristic burst firing following cue or reward presentation, as previously reported, their activity can be explained by the representation of actual movement kinematics. Unlike neighboring pars reticulata GABAergic output neurons, which can represent vector components of position, DA neurons represent vector components of velocity or acceleration. We found neurons related to movements in four directions-up, down, left, right. For horizontal movements, there is significant lateralization of neurons: the left nigra contains more rightward neurons, whereas the right nigra contains more leftward neurons. The relationship between DA activity and movement kinematics was found on both appetitive trials using sucrose and aversive trials using air puff, showing that these neurons belong to a velocity control circuit that can be used for any number of purposes, whether to seek reward or to avoid harm. In support of this conclusion, mimicry of the phasic activation of DA neurons with selective optogenetic stimulation could also generate movements. Contrary to the popular hypothesis that DA neurons encode reward prediction errors, our results suggest that nigrostriatal DA plays an essential role in controlling the kinematics of voluntary movements. We hypothesize that DA signaling implements gain adjustment for adaptive transition control, and describe a new model of the basal ganglia (BG) in which DA functions to adjust the gain of the transition controller. This model has significant implications for our understanding of movement disorders implicating DA and the BG.
Keywords: basal ganglia; dopamine; movement; reward prediction error; striatum; substantia nigra.
Figures
References
-
- Alchian A. A., Allen W. R. (1977). Exchange and Production: Competition, Coordination, and Control. Belmont, CA: Wadsworth Publishing Company.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous