

APP intracellular domain derived from amyloidogenic β- and γ-secretase cleavage regulates neprilysin expression
- PMID: 26074811
- PMCID: PMC4443740
- DOI: 10.3389/fnagi.2015.00077
APP intracellular domain derived from amyloidogenic β- and γ-secretase cleavage regulates neprilysin expression
Abstract
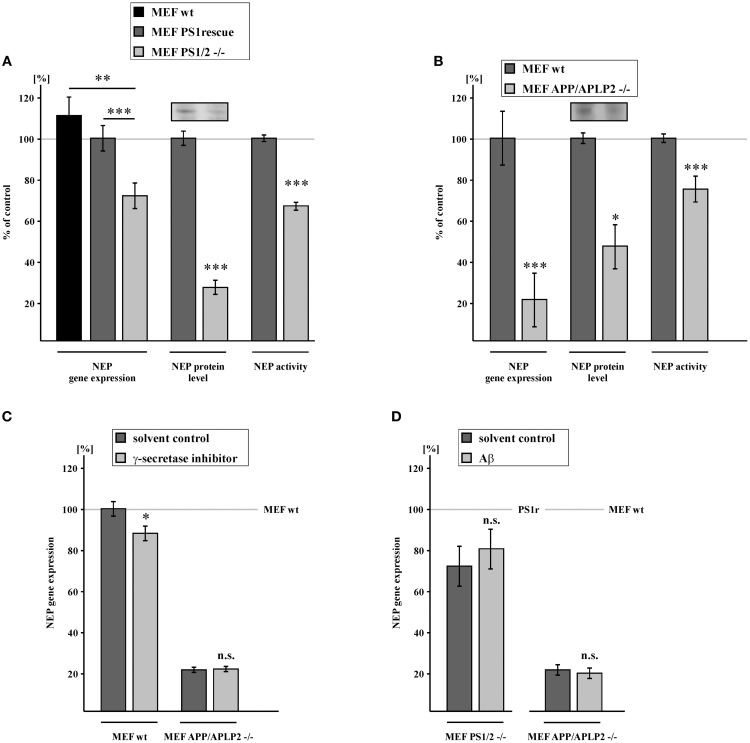
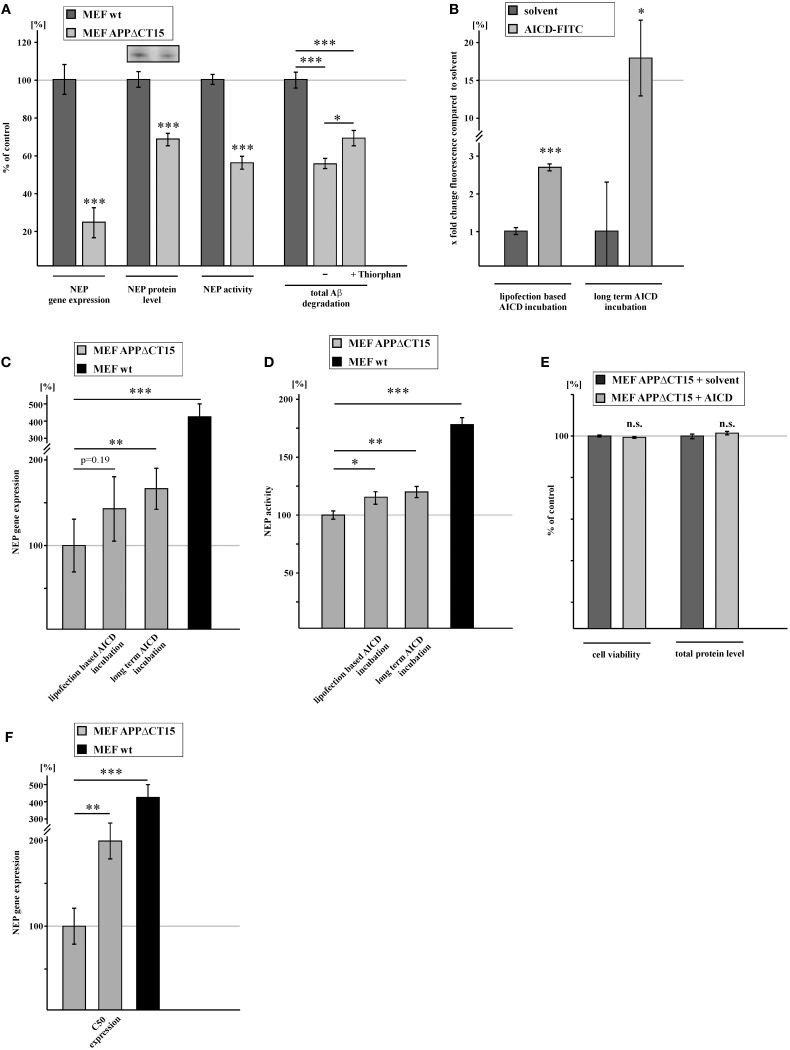
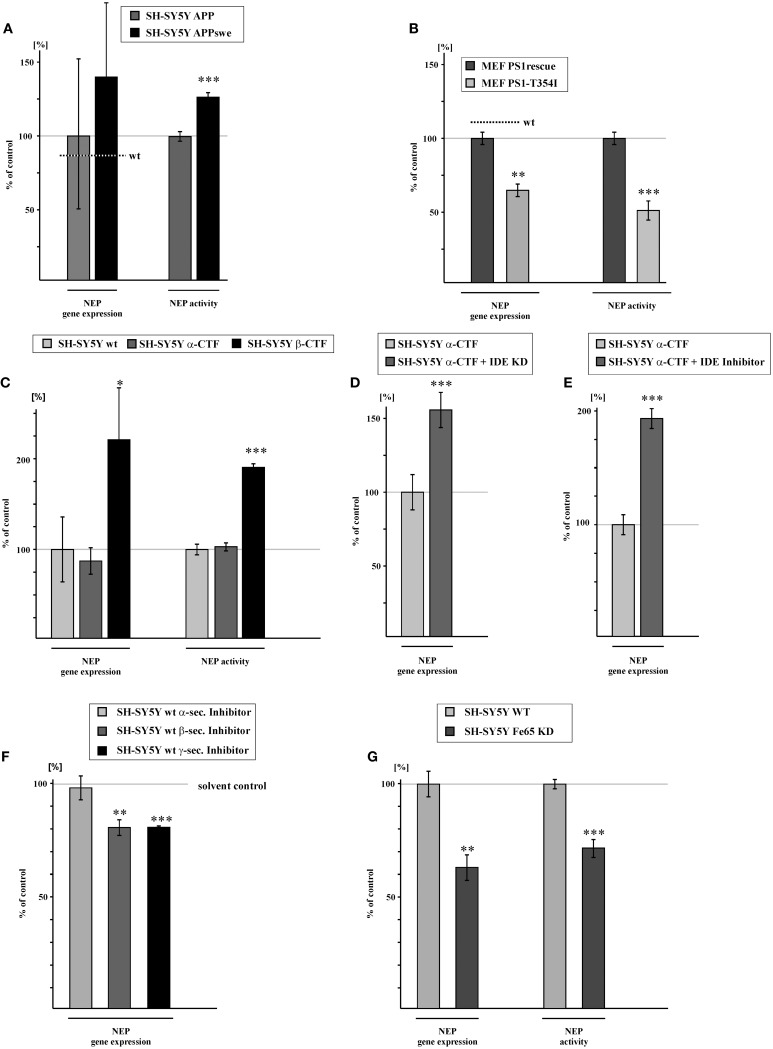
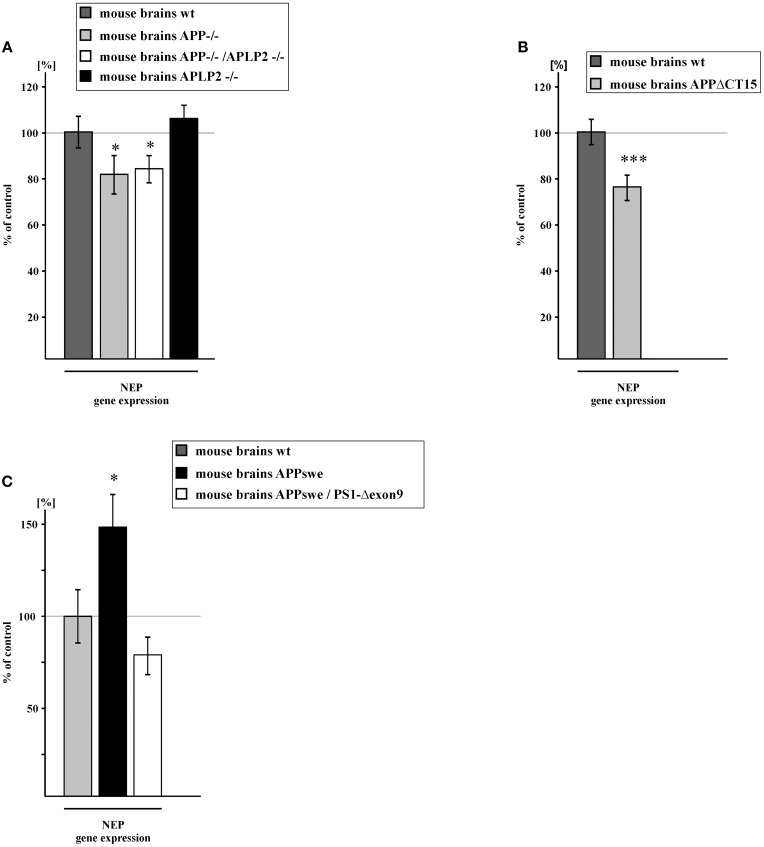
Alzheimer's disease (AD) is characterized by an accumulation of Amyloid-β (Aβ), released by sequential proteolytic processing of the amyloid precursor protein (APP) by β - and γ-secretase. Aβ peptides can aggregate, leading to toxic Aβ oligomers and amyloid plaque formation. Aβ accumulation is not only dependent on de novo synthesis but also on Aβ degradation. Neprilysin (NEP) is one of the major enzymes involved in Aβ degradation. Here we investigate the molecular mechanism of NEP regulation, which is up to now controversially discussed to be affected by APP processing itself. We found that NEP expression is highly dependent on the APP intracellular domain (AICD), released by APP processing. Mouse embryonic fibroblasts devoid of APP processing, either by the lack of the catalytically active subunit of the γ-secretase complex [presenilin (PS) 1/2] or by the lack of APP and the APP-like protein 2 (APLP2), showed a decreased NEP expression, activity and protein level. Similar results were obtained by utilizing cells lacking a functional AICD domain (APPΔCT15) or expressing mutations in the genes encoding for PS1. AICD supplementation or retransfection with an AICD encoding plasmid could rescue the down-regulation of NEP further strengthening the link between AICD and transcriptional NEP regulation, in which Fe65 acts as an important adaptor protein. Especially AICD generated by the amyloidogenic pathway seems to be more involved in the regulation of NEP expression. In line, analysis of NEP gene expression in vivo in six transgenic AD mouse models (APP and APLP2 single knock-outs, APP/APLP2 double knock-out, APP-swedish, APP-swedish/PS1Δexon9, and APPΔCT15) confirmed the results obtained in cell culture. In summary, in the present study we clearly demonstrate an AICD-dependent regulation of the Aβ-degrading enzyme NEP in vitro and in vivo and elucidate the underlying mechanisms that might be beneficial to develop new therapeutic strategies for the treatment of AD.
Keywords: AICD; APP intracellular domain; Alzheimer's disease; Aβ degradation; gene regulation; neprilysin.
Figures
Similar articles
-
Neprilysin and Aβ Clearance: Impact of the APP Intracellular Domain in NEP Regulation and Implications in Alzheimer's Disease.Front Aging Neurosci. 2013 Dec 23;5:98. doi: 10.3389/fnagi.2013.00098. Front Aging Neurosci. 2013. PMID: 24391587 Free PMC article. Review.
-
The transcriptionally active amyloid precursor protein (APP) intracellular domain is preferentially produced from the 695 isoform of APP in a {beta}-secretase-dependent pathway.J Biol Chem. 2010 Dec 31;285(53):41443-54. doi: 10.1074/jbc.M110.141390. Epub 2010 Oct 20. J Biol Chem. 2010. PMID: 20961856 Free PMC article.
-
Regulatory feedback cycle of the insulin-degrading enzyme and the amyloid precursor protein intracellular domain: Implications for Alzheimer's disease.Aging Cell. 2020 Nov;19(11):e13264. doi: 10.1111/acel.13264. Epub 2020 Oct 31. Aging Cell. 2020. PMID: 33128835 Free PMC article.
-
Upregulation of PGC-1α expression by Alzheimer's disease-associated pathway: presenilin 1/amyloid precursor protein (APP)/intracellular domain of APP.Aging Cell. 2014 Apr;13(2):263-72. doi: 10.1111/acel.12183. Epub 2013 Dec 17. Aging Cell. 2014. PMID: 24304563 Free PMC article.
-
AICD nuclear signaling and its possible contribution to Alzheimer's disease.Curr Alzheimer Res. 2012 Feb;9(2):200-16. doi: 10.2174/156720512799361673. Curr Alzheimer Res. 2012. PMID: 21605035 Review.
Cited by
-
Amyloid Precursor Protein (APP) Metabolites APP Intracellular Fragment (AICD), Aβ42, and Tau in Nuclear Roles.J Biol Chem. 2015 Sep 25;290(39):23515-22. doi: 10.1074/jbc.R115.677211. Epub 2015 Aug 21. J Biol Chem. 2015. PMID: 26296890 Free PMC article. Review.
-
Physiological and pathophysiological control of synaptic GluN2B-NMDA receptors by the C-terminal domain of amyloid precursor protein.Elife. 2017 Jul 6;6:e25659. doi: 10.7554/eLife.25659. Elife. 2017. PMID: 28682239 Free PMC article.
-
The Amyloid Cascade Hypothesis 2.0 for Alzheimer's Disease and Aging-Associated Cognitive Decline: From Molecular Basis to Effective Therapy.Int J Mol Sci. 2023 Jul 31;24(15):12246. doi: 10.3390/ijms241512246. Int J Mol Sci. 2023. PMID: 37569624 Free PMC article.
-
Potential Bidirectional Relationship Between Periodontitis and Alzheimer's Disease.Front Physiol. 2020 Jul 3;11:683. doi: 10.3389/fphys.2020.00683. eCollection 2020. Front Physiol. 2020. PMID: 32719612 Free PMC article. Review.
-
Physiological effects of amyloid precursor protein and its derivatives on neural stem cell biology and signaling pathways involved.Neural Regen Res. 2019 Oct;14(10):1661-1671. doi: 10.4103/1673-5374.257511. Neural Regen Res. 2019. PMID: 31169172 Free PMC article.
References
-
- Belyaev N. D., Kellett K. A., Beckett C., Makova N. Z., Revett T. J., Nalivaeva N. N., et al. (2010). The transcriptionally active amyloid precursor protein (APP) intracellular domain is preferentially produced from the 695 isoform of APP in a {beta}-secretase-dependent pathway. J. Biol. Chem. 285, 41443–41454. 10.1074/jbc.M110.141390 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources