

Redundancy between Cysteine Cathepsins in Murine Experimental Autoimmune Encephalomyelitis
- PMID: 26075905
- PMCID: PMC4468166
- DOI: 10.1371/journal.pone.0128945
Redundancy between Cysteine Cathepsins in Murine Experimental Autoimmune Encephalomyelitis
Abstract
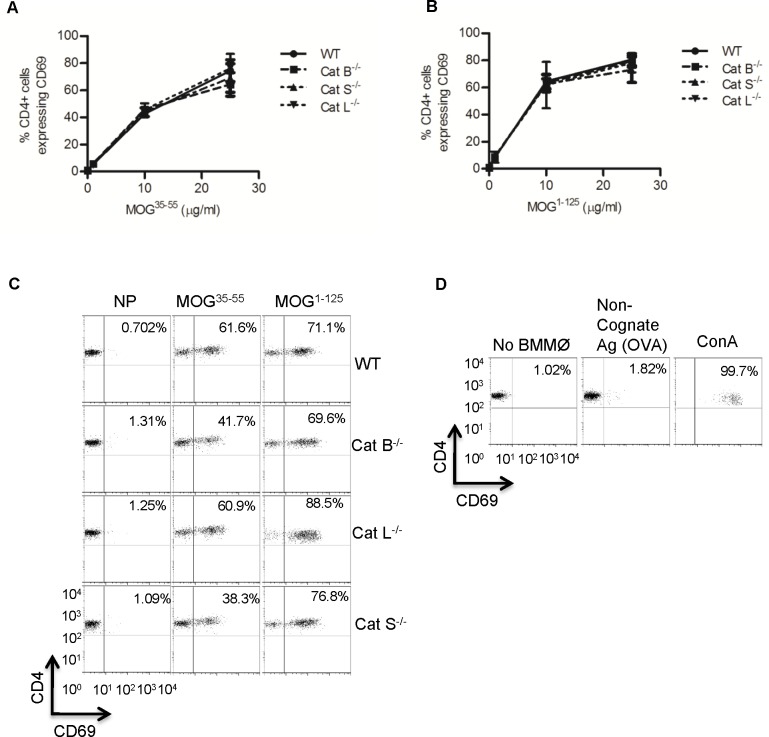
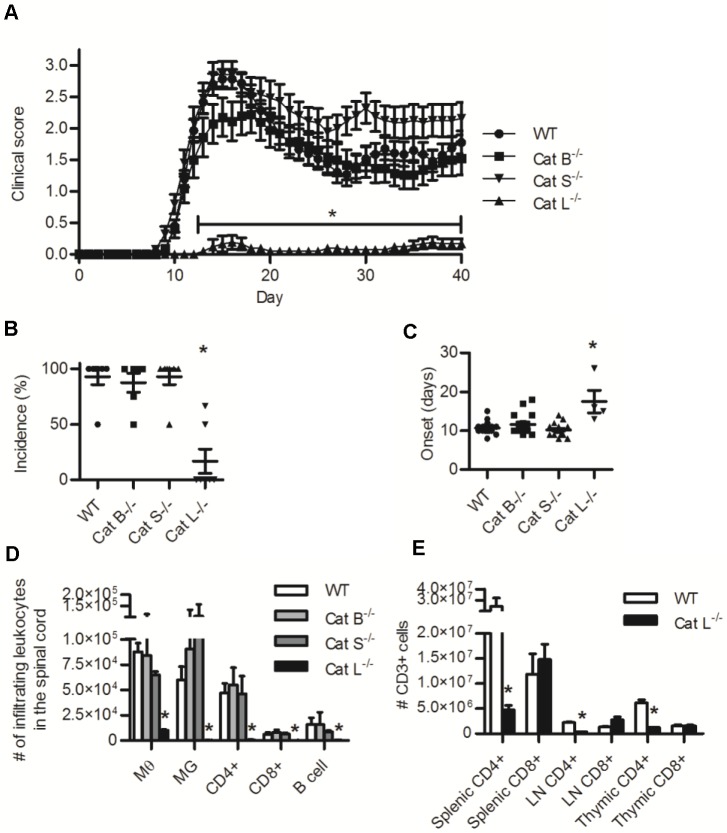
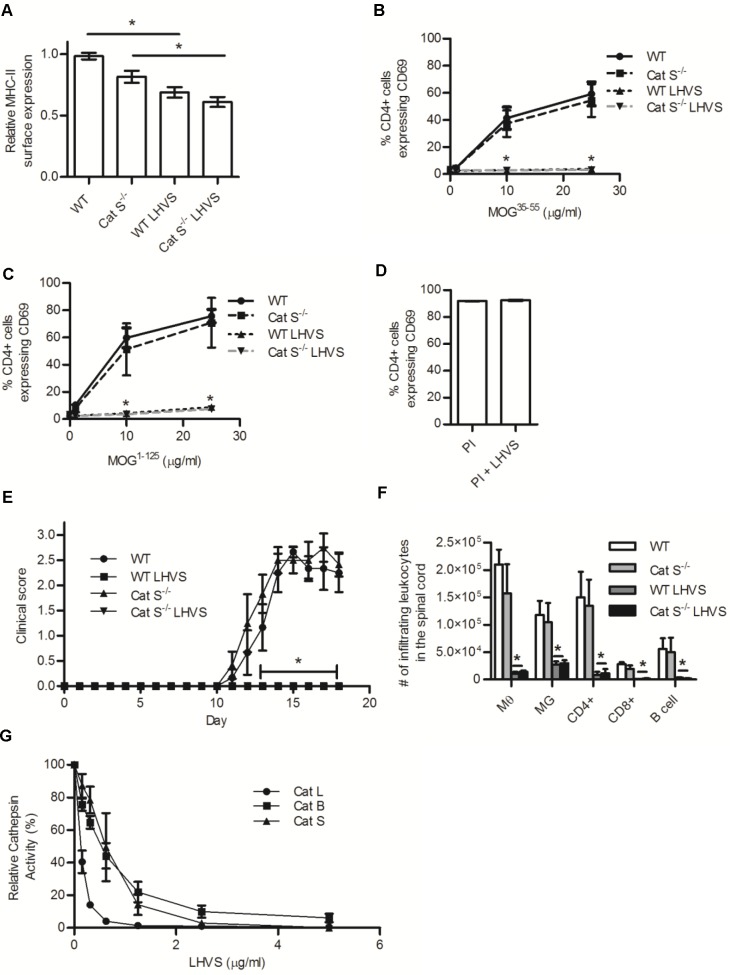
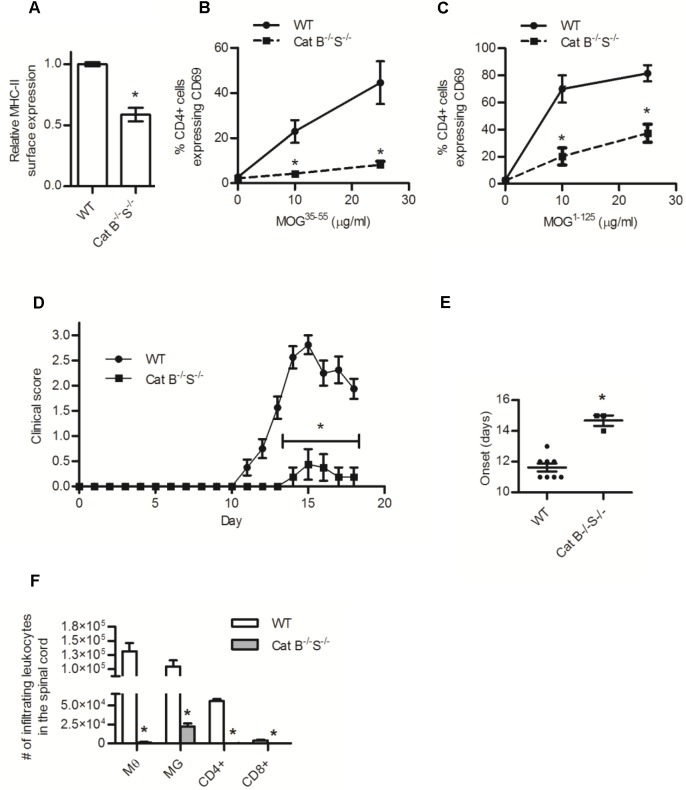
The cysteine cathepsins B, S, and L are functionally linked to antigen processing, and hence to autoimmune disorders such as multiple sclerosis. Stemming from several studies that demonstrate that mice can be protected from experimental autoimmune encephalomyelitis (EAE) through the pharmacologic inhibition of cysteine cathepsins, it has been suggested that targeting these enzymes in multiple sclerosis may be of therapeutic benefit. Utilizing mice deficient in cysteine cathepsins both individually and in combination, we found that the myelin-associated antigen myelin oligodendrocyte glycoprotein (MOG) was efficiently processed and presented by macrophages to CD4+ T cells in the individual absence of cathepsin B, S or L. Similarly, mice deficient in cathepsin B or S were susceptible to MOG-induced EAE and displayed clinical progression and immune infiltration into the CNS, similar to their wild-type counterparts. Owing to a previously described CD4+ T cell deficiency in mice deficient in cathepsin L, such mice were protected from EAE. When multiple cysteine cathepsins were simultaneously inhibited via genetic deletion of both cathepsins B and S, or by a cathepsin inhibitor (LHVS), MHC-II surface expression, MOG antigen presentation and EAE were attenuated or prevented. This study demonstrates the functional redundancy between cathepsin B, S and L in EAE, and suggests that the inhibition of multiple cysteine cathepsins may be needed to modulate autoimmune disorders such as multiple sclerosis.
Conflict of interest statement
Figures
References
-
- Vasiljeva O, Reinheckel T, Peters C, Turk D, Turk V, Turk B. Emerging roles of cysteine cathepsins in disease and their potential as drug targets. Curr Pharm Des. 2007;13(4):387–403. Epub 2007/02/22. . - PubMed
-
- Honey K, Duff M, Beers C, Brissette WH, Elliott EA, Peters C, et al. Cathepsin S regulates the expression of cathepsin L and the turnover of gamma-interferon-inducible lysosomal thiol reductase in B lymphocytes. J Biol Chem. 2001;276(25):22573–8. Epub 2001/04/18. 10.1074/jbc.M101851200M101851200 [pii]. . - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
