

Derivation of Cinnamon Blocks Leukocyte Attachment by Interacting with Sialosides
- PMID: 26076445
- PMCID: PMC4468131
- DOI: 10.1371/journal.pone.0130389
Derivation of Cinnamon Blocks Leukocyte Attachment by Interacting with Sialosides
Abstract
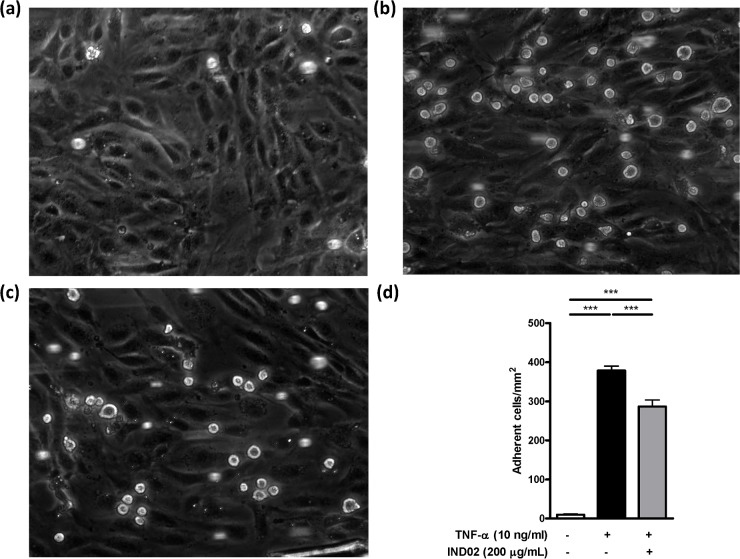
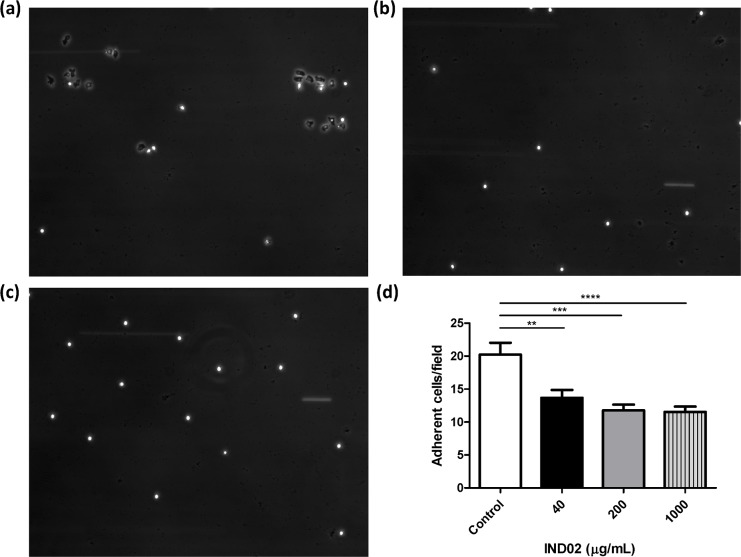
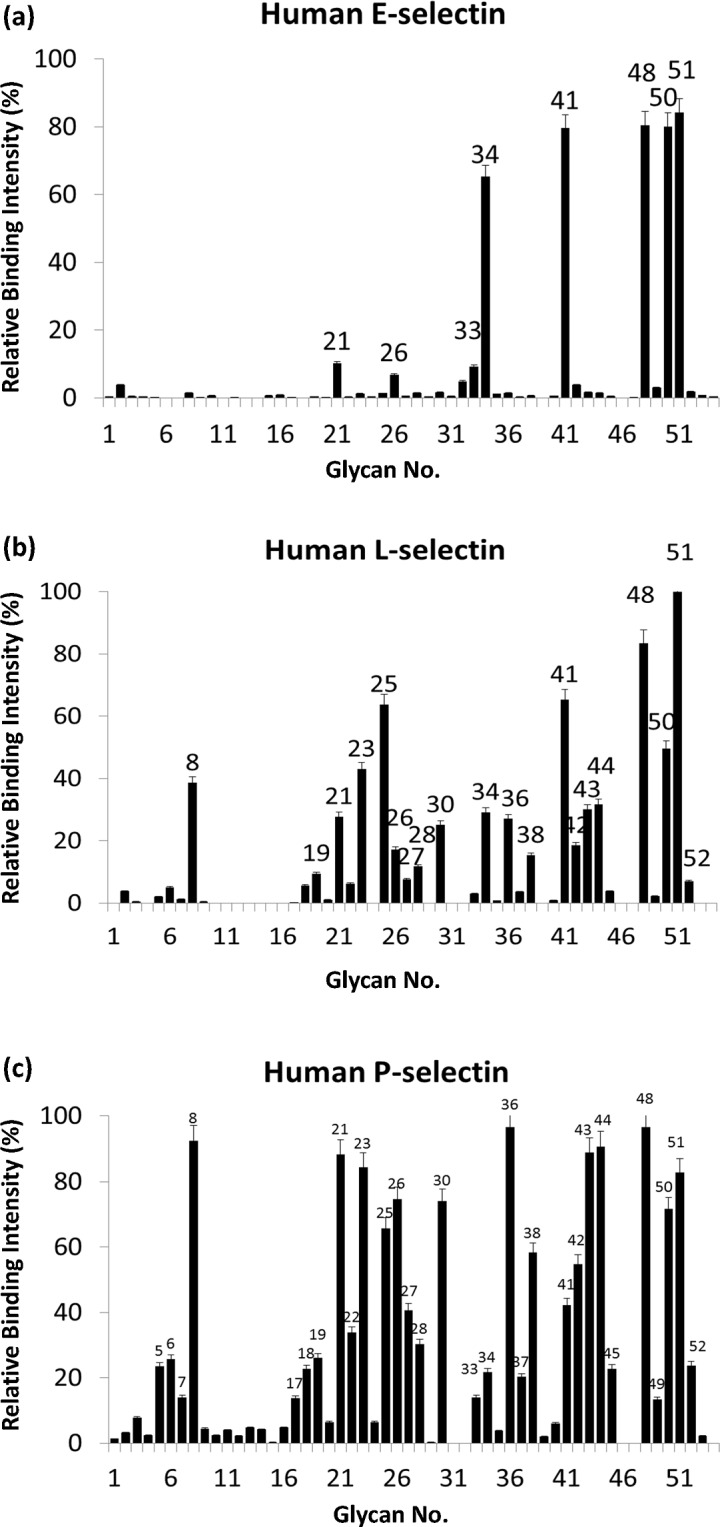
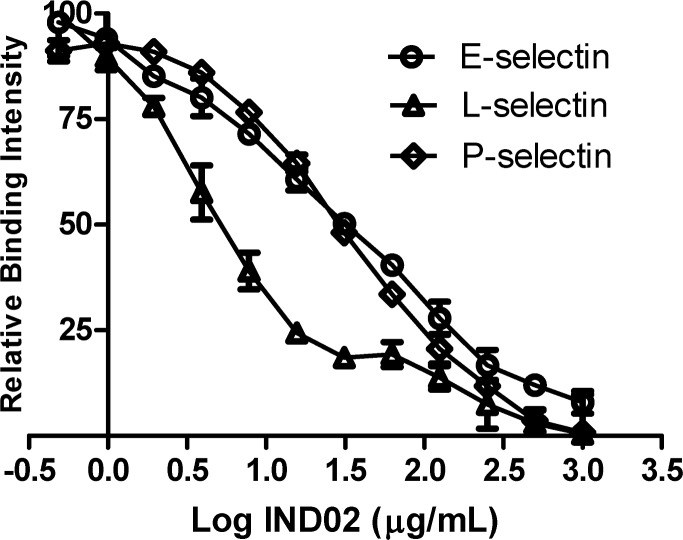
Molecules derived from cinnamon have demonstrated diverse pharmacological activities against infectious pathogens, diabetes and inflammatory diseases. This study aims to evaluate the effect of the cinnamon-derived molecule IND02 on the adhesion of leukocytes to host cells. The anti-inflammatory ability of IND02, a pentameric procyanidin type A polyphenol polymer isolated from cinnamon alcohol extract, was examined. Pretreatment with IND02 significantly reduced the attachment of THP-1 cells or neutrophils to TNF-α-activated HUVECs or E-selectin/ICAM-1, respectively. IND02 also reduced the binding of E-, L- and P-selectins with sialosides. Furthermore, IND02 could agglutinate human red blood cells (RBC), and the agglutination could be disrupted by sialylated glycoprotein. Our findings demonstrate that IND02, a cinnamon-derived compound, can interact with sialosides and block the binding of selectins and leukocytes with sialic acids.
Conflict of interest statement
Figures
References
-
- Koppikar SJ, Choudhari AS, Suryavanshi SA, Kumari S, Chattopadhyay S, Kaul-Ghanekar R. Aqueous cinnamon extract (ACE-c) from the bark of Cinnamomum cassia causes apoptosis in human cervical cancer cell line (SiHa) through loss of mitochondrial membrane potential. BMC Cancer. 2010;10: 210 10.1186/1471-2407-10-210 - DOI - PMC - PubMed
-
- Shan B, Cai YZ, Brooks JD, Corke H. Antibacterial properties and major bioactive components of cinnamon stick (Cinnamomum burmannii): activity against foodborne pathogenic bacteria. J Agric Food Chem. 2007;55: 5484–5490. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
