

The sodium-activated potassium channel Slack is required for optimal cognitive flexibility in mice
- PMID: 26077685
- PMCID: PMC4478330
- DOI: 10.1101/lm.037820.114
The sodium-activated potassium channel Slack is required for optimal cognitive flexibility in mice
Abstract
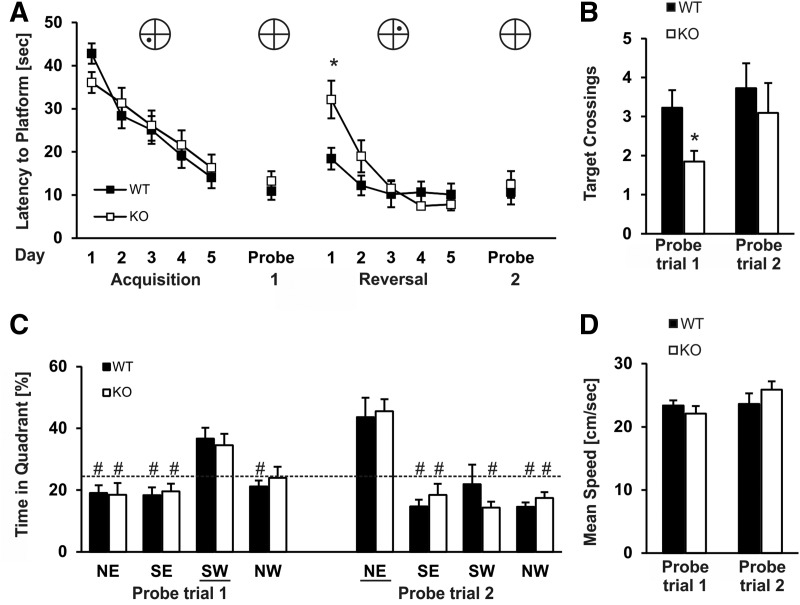
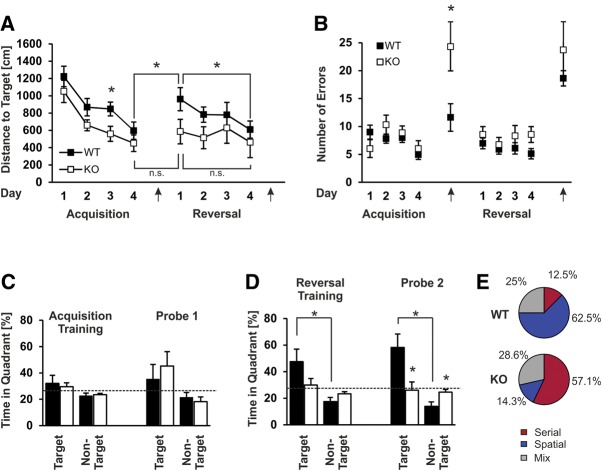
Kcnt1 encoded sodium-activated potassium channels (Slack channels) are highly expressed throughout the brain where they modulate the firing patterns and general excitability of many types of neurons. Increasing evidence suggests that Slack channels may be important for higher brain functions such as cognition and normal intellectual development. In particular, recent findings have shown that human Slack mutations produce very severe intellectual disability and that Slack channels interact directly with the Fragile X mental retardation protein (FMRP), a protein that when missing or mutated results in Fragile X syndrome (FXS), the most common form of inherited intellectual disability and autism in humans. We have now analyzed a recently developed Kcnt1 null mouse model in several behavioral tasks to assess which aspects of memory and learning are dependent on Slack. We demonstrate that Slack deficiency results in mildly altered general locomotor activity, but normal working memory, reference memory, as well as cerebellar control of motor functions. In contrast, we find that Slack channels are required for cognitive flexibility, including reversal learning processes and the ability to adapt quickly to unfamiliar situations and environments. Our data reveal that hippocampal-dependent spatial learning capabilities require the proper function of Slack channels.
© 2015 Bausch et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
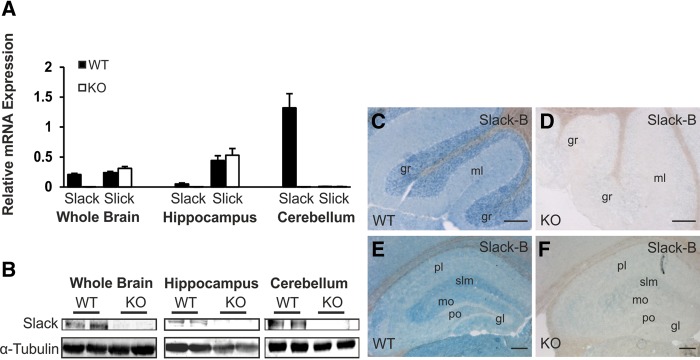
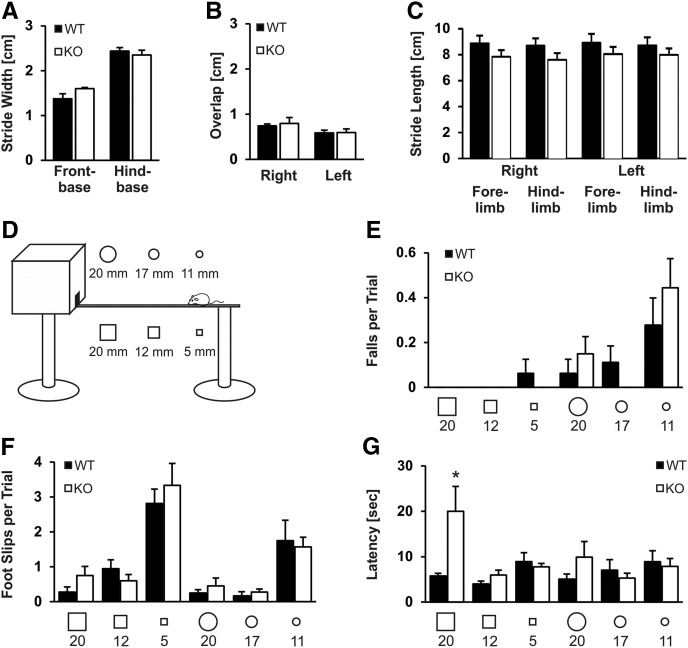
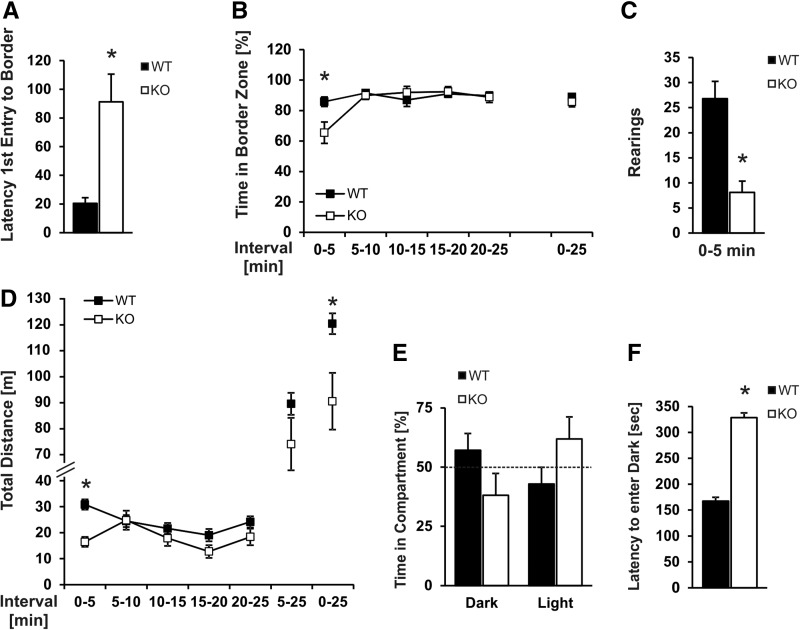
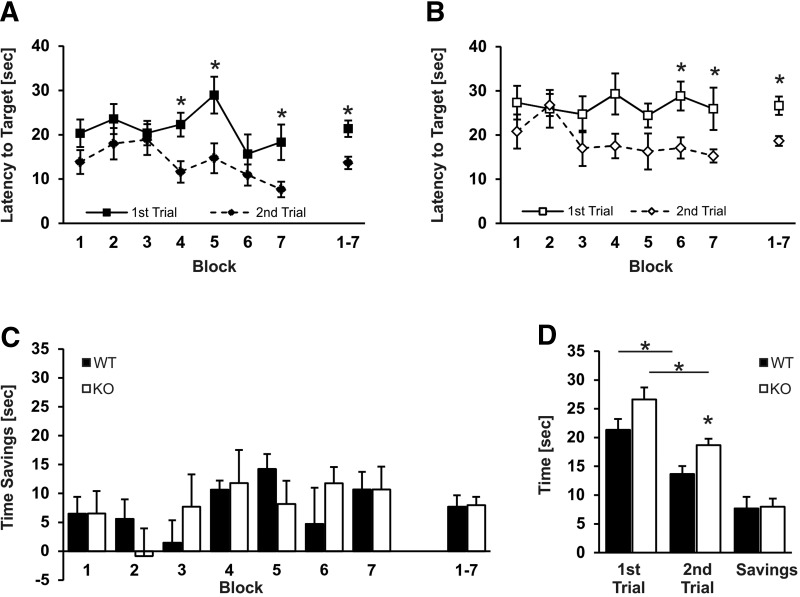
References
-
- Aminkeng F. 2012. KCNT1 mutations in ADNFLE and MMPSI: a new driver in the etiology and pathophysiology of early-onset epileptic syndromes. Clin Genet 83: 319–320. - PubMed
-
- Baker KB, Wray SP, Ritter R, Mason S, Lanthorn TH, Savelieva KV. 2010. Male and female Fmr1 knockout mice on C57 albino background exhibit spatial learning and memory impairments. Genes Brain Behav 9: 562–574. - PubMed
-
- Bakker CE, Oostra BA. 2003. Understanding fragile X syndrome: insights from animal models. Cytogenet Genome Res 100: 111–123. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases