

The lysophosphatidylserine receptor GPR174 constrains regulatory T cell development and function
- PMID: 26077720
- PMCID: PMC4493414
- DOI: 10.1084/jem.20141827
The lysophosphatidylserine receptor GPR174 constrains regulatory T cell development and function
Abstract
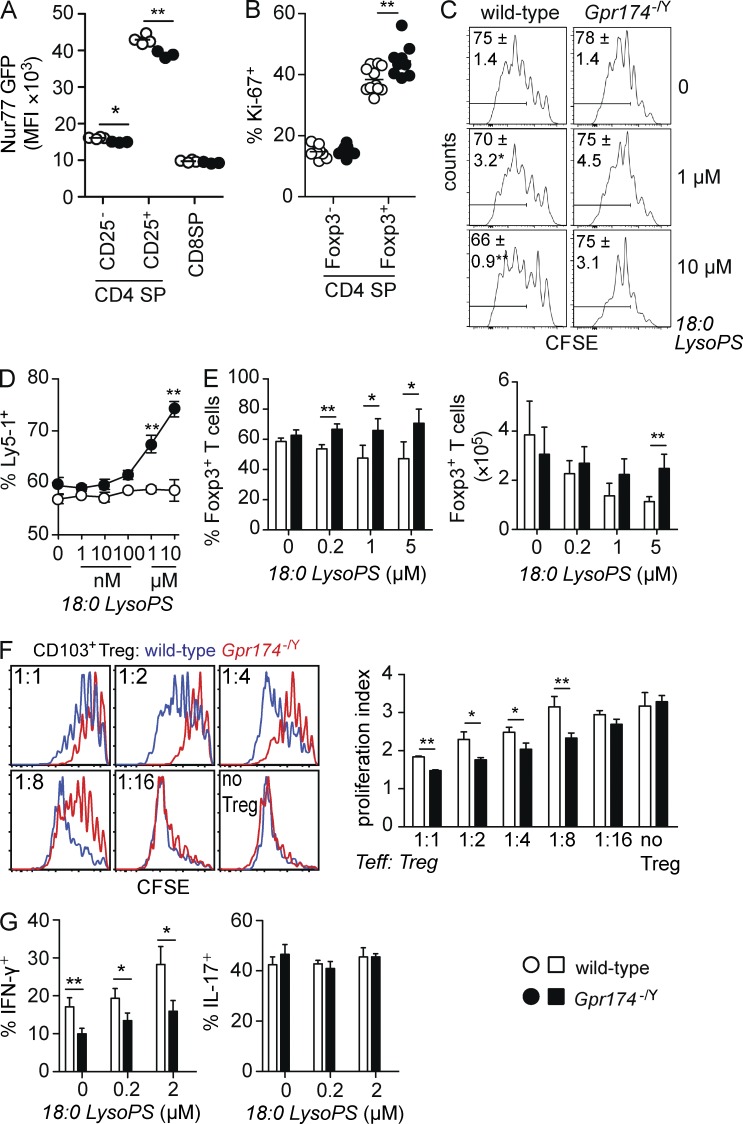
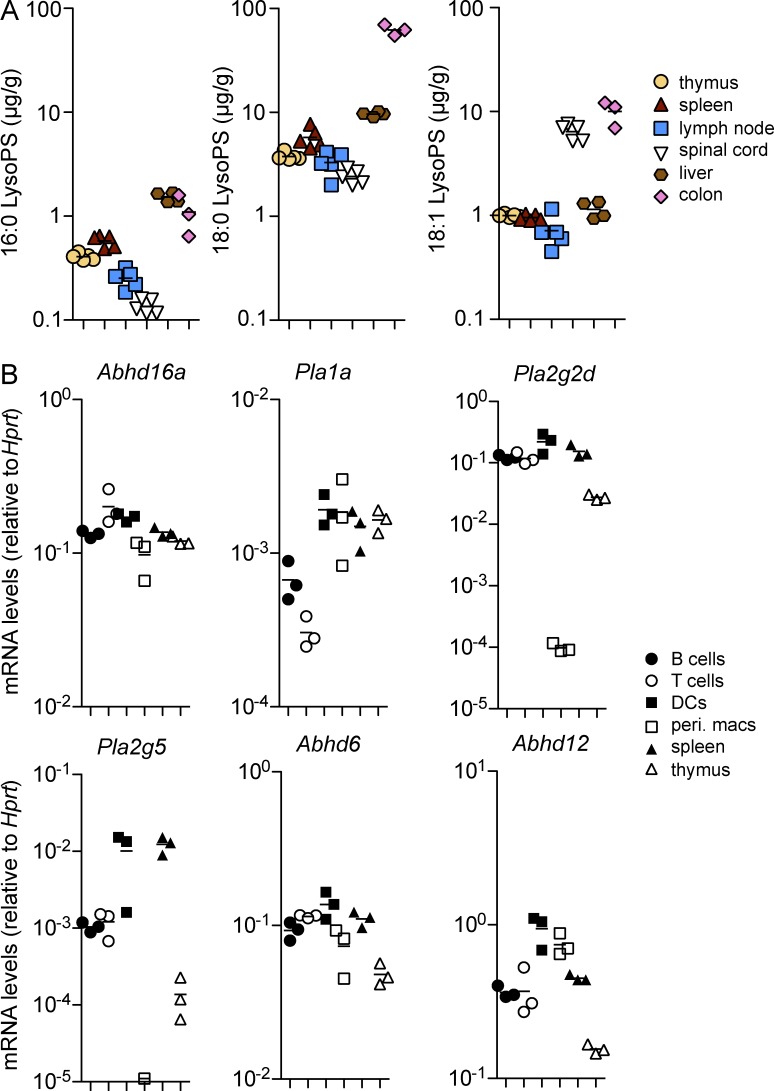
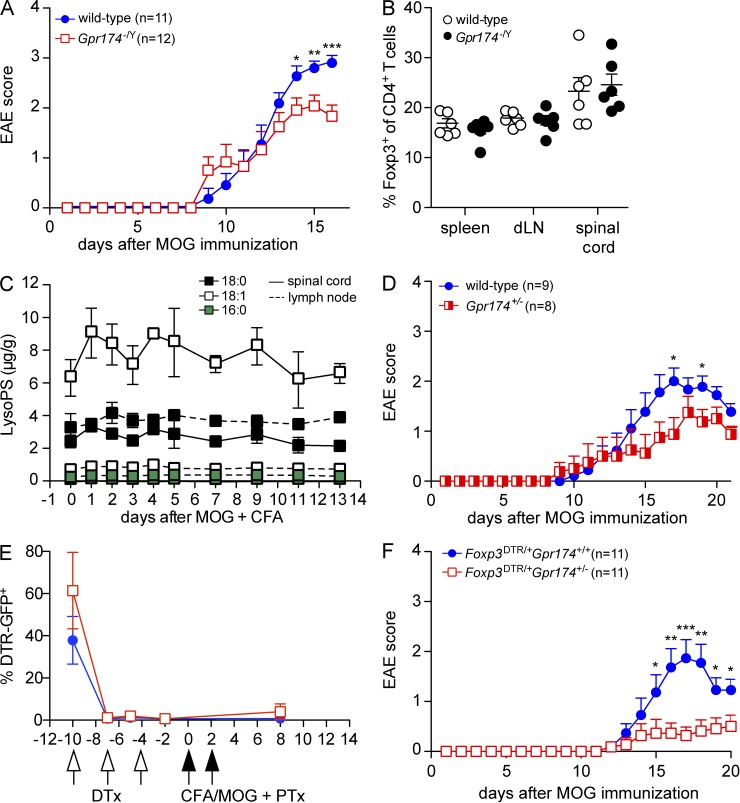
Regulatory T cell (T reg cell) numbers and activities are tightly calibrated to maintain immune homeostasis, but the mechanisms involved are incompletely defined. Here, we report that the lysophosphatidylserine (LysoPS) receptor GPR174 is abundantly expressed in developing and mature T reg cells. In mice that lacked this X-linked gene, T reg cell generation in the thymus was intrinsically favored, and a higher fraction of peripheral T reg cells expressed CD103. LysoPS could act in vitro via GPR174 to suppress T cell proliferation and T reg cell generation. In vivo, LysoPS was detected in lymphoid organ and spinal cord tissues and was abundant in the colon. Gpr174(-/Y) mice were less susceptible to experimental autoimmune encephalomyelitis than wild-type mice, and GPR174 deficiency in T reg cells contributed to this phenotype. This study provides evidence that a bioactive lipid, LysoPS, negatively influences T reg cell accumulation and activity through GPR174. As such, GPR174 antagonists might have therapeutic potential for promoting immune regulation in the context of autoimmune disease.
© 2015 Barnes et al.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
