

Orientation Maps in V1 and Non-Euclidean Geometry
- PMID: 26082007
- PMCID: PMC4469697
- DOI: 10.1186/s13408-015-0024-7
Orientation Maps in V1 and Non-Euclidean Geometry
Abstract
In the primary visual cortex, the processing of information uses the distribution of orientations in the visual input: neurons react to some orientations in the stimulus more than to others. In many species, orientation preference is mapped in a remarkable way on the cortical surface, and this organization of the neural population seems to be important for visual processing. Now, existing models for the geometry and development of orientation preference maps in higher mammals make a crucial use of symmetry considerations. In this paper, we consider probabilistic models for V1 maps from the point of view of group theory; we focus on Gaussian random fields with symmetry properties and review the probabilistic arguments that allow one to estimate pinwheel densities and predict the observed value of π. Then, in order to test the relevance of general symmetry arguments and to introduce methods which could be of use in modeling curved regions, we reconsider this model in the light of group representation theory, the canonical mathematics of symmetry. We show that through the Plancherel decomposition of the space of complex-valued maps on the Euclidean plane, each infinite-dimensional irreducible unitary representation of the special Euclidean group yields a unique V1-like map, and we use representation theory as a symmetry-based toolbox to build orientation maps adapted to the most famous non-Euclidean geometries, viz. spherical and hyperbolic geometry. We find that most of the dominant traits of V1 maps are preserved in these; we also study the link between symmetry and the statistics of singularities in orientation maps, and show what the striking quantitative characteristics observed in animals become in our curved models.
Figures



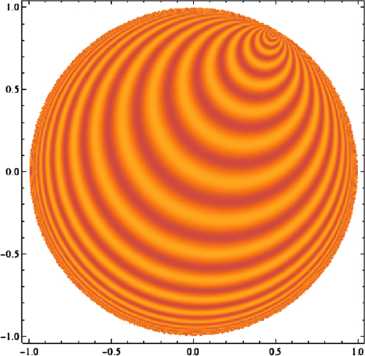

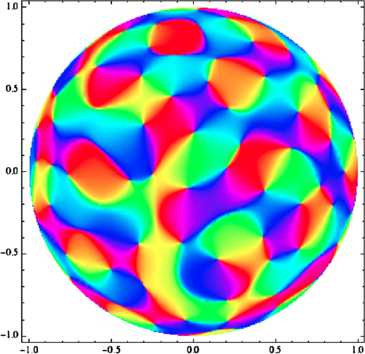
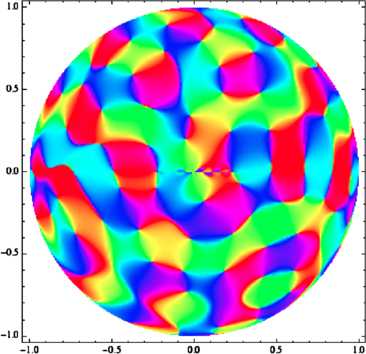
References
-
- De Angelis GC, Ohzawa I, Freeman RD. Spatiotemporal organization of simple-cell receptive fields in the cat’s striate cortex. I. General characteristics and postnatal development. J Neurophysiol. 1993;69:1091–1117. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
