

LABA1, a Domestication Gene Associated with Long, Barbed Awns in Wild Rice
- PMID: 26082172
- PMCID: PMC4531357
- DOI: 10.1105/tpc.15.00260
LABA1, a Domestication Gene Associated with Long, Barbed Awns in Wild Rice
Abstract
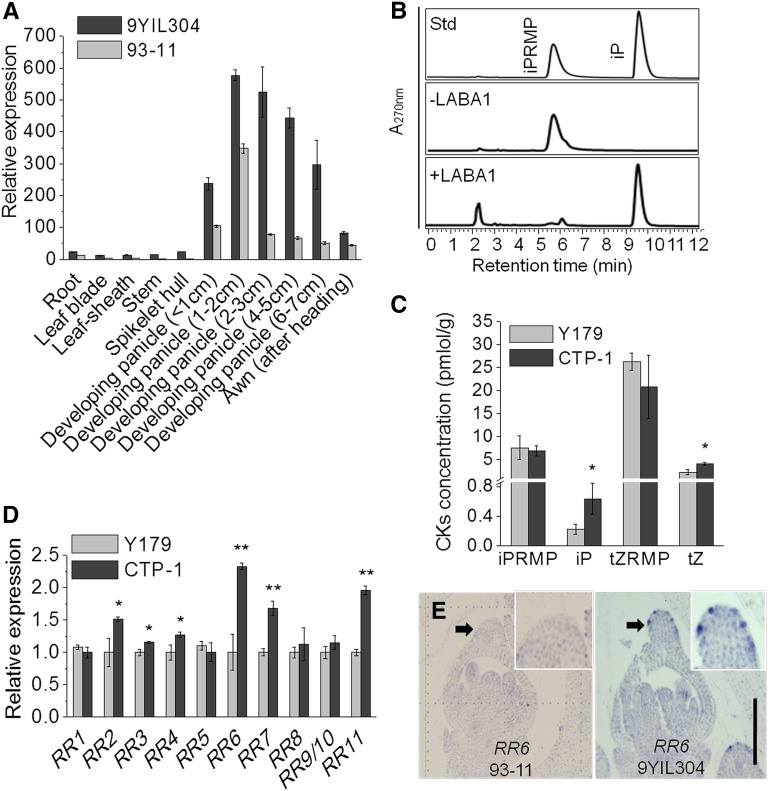
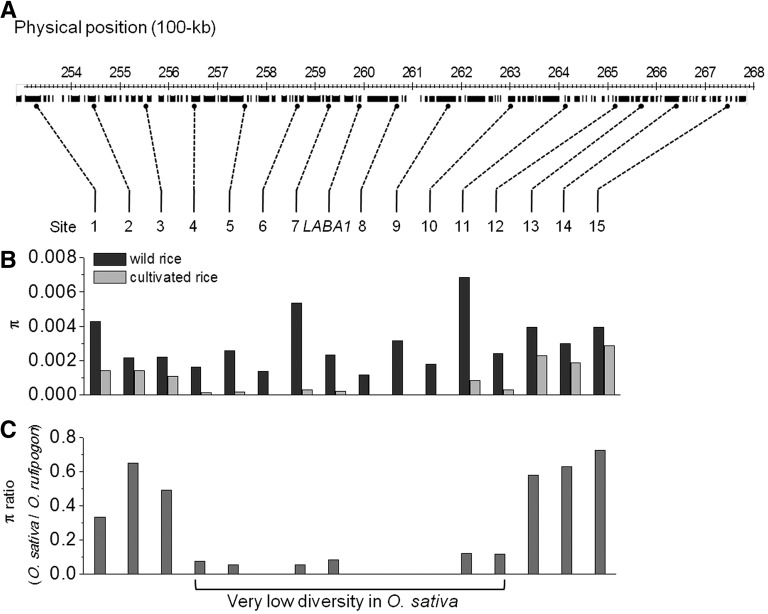
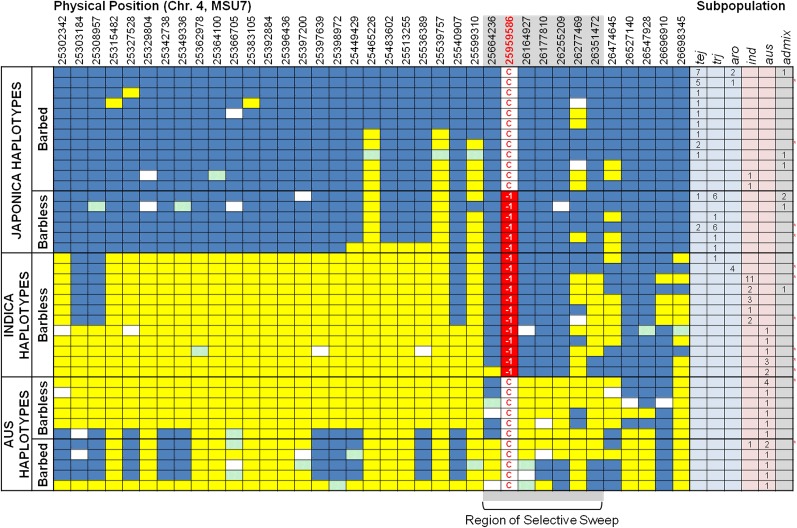
Common wild rice (Oryza rufipogon), the wild relative of Asian cultivated rice (Oryza sativa), flaunts long, barbed awns, which are necessary for efficient propagation and dissemination of seeds. By contrast, O. sativa cultivars have been selected to be awnless or to harbor short, barbless awns, which facilitate seed processing and storage. The transition from long, barbed awns to short, barbless awns was a crucial event in rice domestication. Here, we show that the presence of long, barbed awns in wild rice is controlled by a major gene on chromosome 4, LONG AND BARBED AWN1 (LABA1), which encodes a cytokinin-activating enzyme. A frame-shift deletion in LABA1 of cultivated rice reduces the cytokinin concentration in awn primordia, disrupting barb formation and awn elongation. Sequencing analysis demonstrated low nucleotide diversity and a selective sweep encompassing an ∼800-kb region around the derived laba1 allele in cultivated rice. Haplotype analysis revealed that the laba1 allele originated in the japonica subspecies and moved into the indica gene pool via introgression, suggesting that humans selected for this locus in early rice domestication. Identification of LABA1 provides new insights into rice domestication and also sheds light on the molecular mechanism underlying awn development.
© 2015 American Society of Plant Biologists. All rights reserved.
Figures




Comment in
-
Domesticated versus Wild Rice? Bring It Awn!Plant Cell. 2015 Jul;27(7):1818. doi: 10.1105/tpc.15.00504. Epub 2015 Jun 16. Plant Cell. 2015. PMID: 26082171 Free PMC article. No abstract available.
References
-
- Dewitte W., Scofield S., Alcasabas A.A., Maughan S.C., Menges M., Braun N., Collins C., Nieuwland J., Prinsen E., Sundaresan V., Murray J.A. (2007). Arabidopsis CYCD3 D-type cyclins link cell proliferation and endocycles and are rate-limiting for cytokinin responses. Proc. Natl. Acad. Sci. USA 104: 14537–14542. - PMC - PubMed
-
- Elbaum R., Zaltzman L., Burgert I., Fratzl P. (2007). The role of wheat awns in the seed dispersal unit. Science 316: 884–886. - PubMed
-
- Fan C., Yu S., Wang C., Xing Y. (2009). A causal C-A mutation in the second exon of GS3 highly associated with rice grain length and validated as a functional marker. Theor. Appl. Genet. 118: 465–472. - PubMed
-
- Fu Q., Zhang P., Tan L., Zhu Z., Ma D., Fu Y., Zhan X., Cai H., Sun C. (2010). Analysis of QTLs for yield-related traits in Yuanjiang common wild rice (Oryza rufipogon Griff.). J. Genet. Genomics 37: 147–157. - PubMed
-
- Fuller D., Sato Y., Castillo C. (2010). Consilience of genetics and archaeobotany in the entangled history of rice. Archaeol. Anthropol. Sci. 2: 115–131.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
