

The FAD-dependent glycerol-3-phosphate dehydrogenase of Giardia duodenalis: an unconventional enzyme that interacts with the g14-3-3 and it is a target of the antitumoral compound NBDHEX
- PMID: 26082764
- PMCID: PMC4450592
- DOI: 10.3389/fmicb.2015.00544
The FAD-dependent glycerol-3-phosphate dehydrogenase of Giardia duodenalis: an unconventional enzyme that interacts with the g14-3-3 and it is a target of the antitumoral compound NBDHEX
Abstract
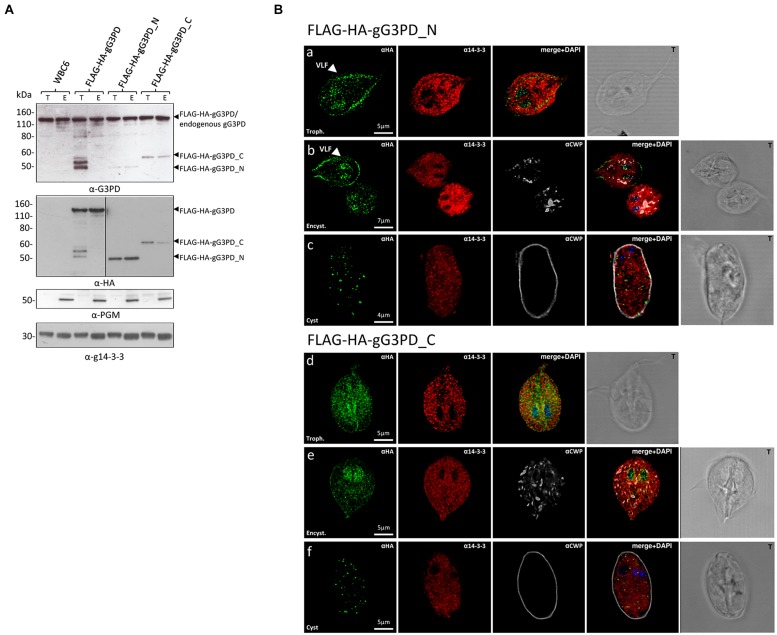
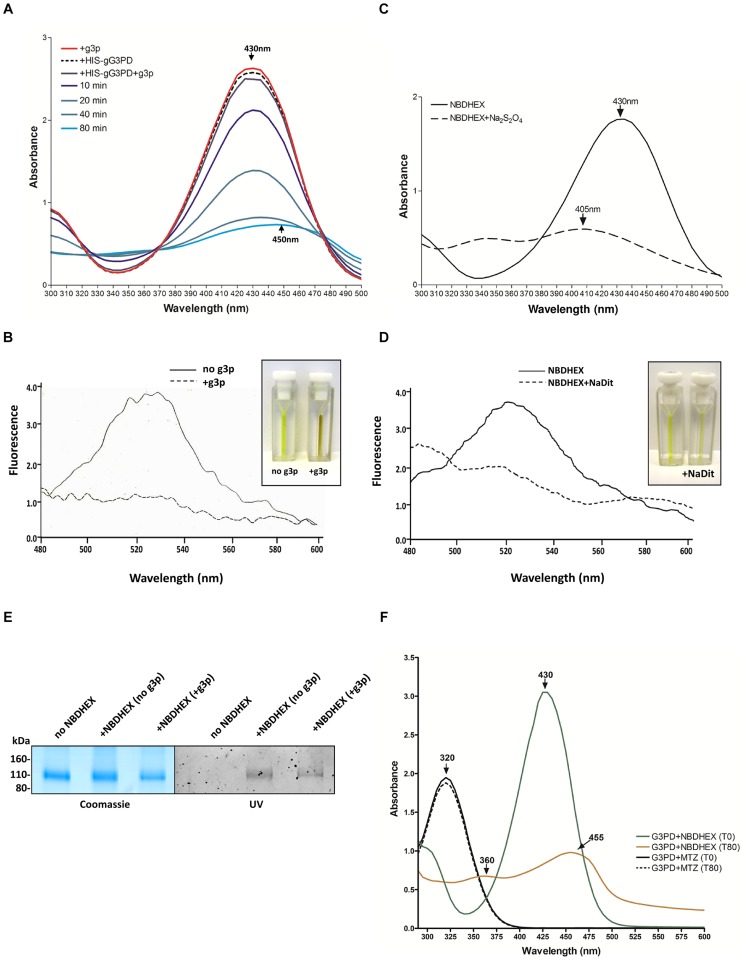
The flagellated protozoan Giardia duodenalis is a worldwide parasite causing giardiasis, an acute and chronic diarrheal disease. Metabolism in G. duodenalis has a limited complexity thus making metabolic enzymes ideal targets for drug development. However, only few metabolic pathways (i.e., carbohydrates) have been described so far. Recently, the parasite homolog of the mitochondrial-like glycerol-3-phosphate dehydrogenase (gG3PD) has been identified among the interactors of the g14-3-3 protein. G3PD is involved in glycolysis, electron transport, glycerophospholipids metabolism, and hyperosmotic stress response, and is emerging as promising target in tumor treatment. In this work, we demonstrate that gG3PD is a functional flavoenzyme able to convert glycerol-3-phosphate into dihydroxyacetone phosphate and that its activity and the intracellular glycerol level increase during encystation. Taking advantage of co-immunoprecipitation assays and deletion mutants, we provide evidence that gG3PD and g14-3-3 interact at the trophozoite stage, the intracellular localization of gG3PD is stage dependent and it partially co-localizes with mitosomes during cyst development. Finally, we demonstrate that the gG3PD activity is affected by the antitumoral compound 6-(7-nitro-2,1,3-benzoxadiazol-4-ylthio)hexanol, that results more effective in vitro at killing G. duodenalis trophozoites than the reference drug metronidazole. Overall, our results highlight the involvement of gG3PD in processes crucial for the parasite survival thus proposing this enzyme as target for novel antigiardial interventions.
Keywords: 14-3-3 protein; FAD-dependent glycerol-3-phoshate dehydrogenase; Giardia duodenalis; NBDHEX; encystation; energy metabolism; mitosome; nitroreduction.
Figures





References
-
- Albertyn J., Hohmann S., Thevelein J. M., Prior B. A. (1994). GPD1 which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol. Cell. Biol. 14 4135–4144. - PMC - PubMed
-
- Andersson J. O., Sjögren A. M., Horner D. S., Murphy C. A., Dyal P. L., Svärd S. G., et al. (2007). A genomic survey of the fish parasite Spironucleus salmonicida indicates genomic plasticity among diplomonads and significant lateral gene transfer in eukaryote genome evolution. BMC Genomics 8:51 10.1186/1471-2164-8-51 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials