

Interleukin-33 in Tissue Homeostasis, Injury, and Inflammation
- PMID: 26084021
- PMCID: PMC4471869
- DOI: 10.1016/j.immuni.2015.06.006
Interleukin-33 in Tissue Homeostasis, Injury, and Inflammation
Abstract
Interleukin-33 (IL-33) is a nuclear-associated cytokine of the IL-1 family originally described as a potent inducer of allergic type 2 immunity. IL-33 signals via the receptor ST2, which is highly expressed on group 2 innate lymphoid cells (ILC2s) and T helper 2 (Th2) cells, thus underpinning its association with helminth infection and allergic pathology. Recent studies have revealed ST2 expression on subsets of regulatory T cells, and for a role for IL-33 in tissue homeostasis and repair that suggests previously unrecognized interactions within these cellular networks. IL-33 can participate in pathologic fibrotic reactions, or, in the setting of microbial invasion, can cooperate with inflammatory cytokines to promote responses by cytotoxic NK cells, Th1 cells, and CD8(+) T cells. Here, we highlight the regulation and function of IL-33 and ST2 and review their roles in homeostasis, damage, and inflammation, suggesting a conceptual framework for future studies.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no conflicting financial interests.
Figures
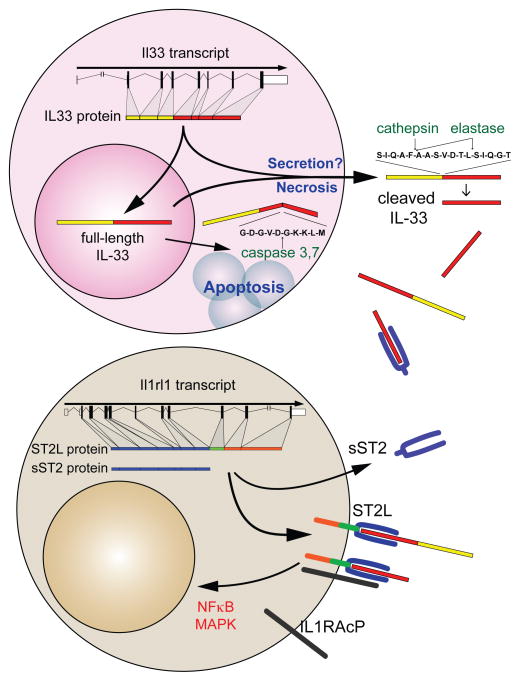
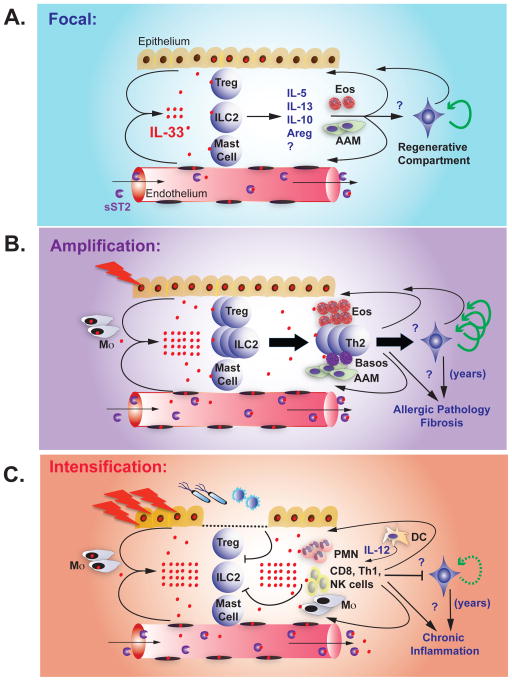
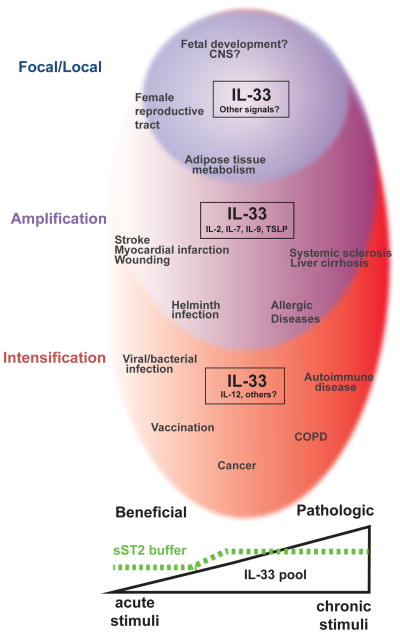
References
-
- Ali S, Mohs A, Thomas M, Klare J, Ross R, Schmitz ML, Martin MU. The dual function cytokine IL-33 interacts with the transcription factor NF-κB to dampen NF-κB-stimulated gene transcription. J Immunol. 2011;187:1609–1616. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
