

Complement Regulates Nutrient Influx and Metabolic Reprogramming during Th1 Cell Responses
- PMID: 26084023
- PMCID: PMC4518498
- DOI: 10.1016/j.immuni.2015.05.024
Complement Regulates Nutrient Influx and Metabolic Reprogramming during Th1 Cell Responses
Abstract
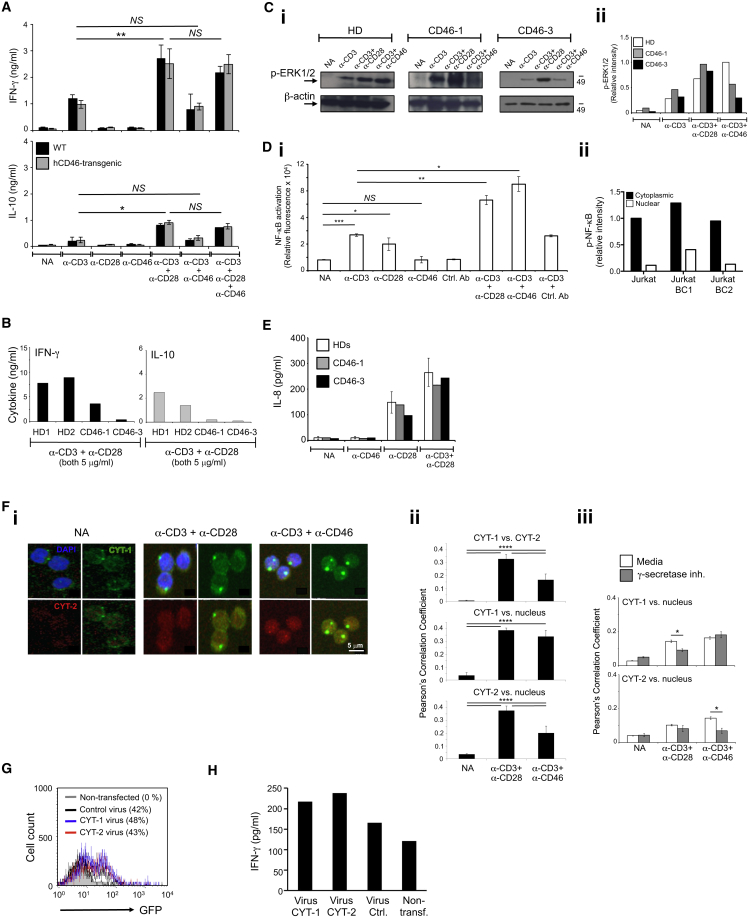
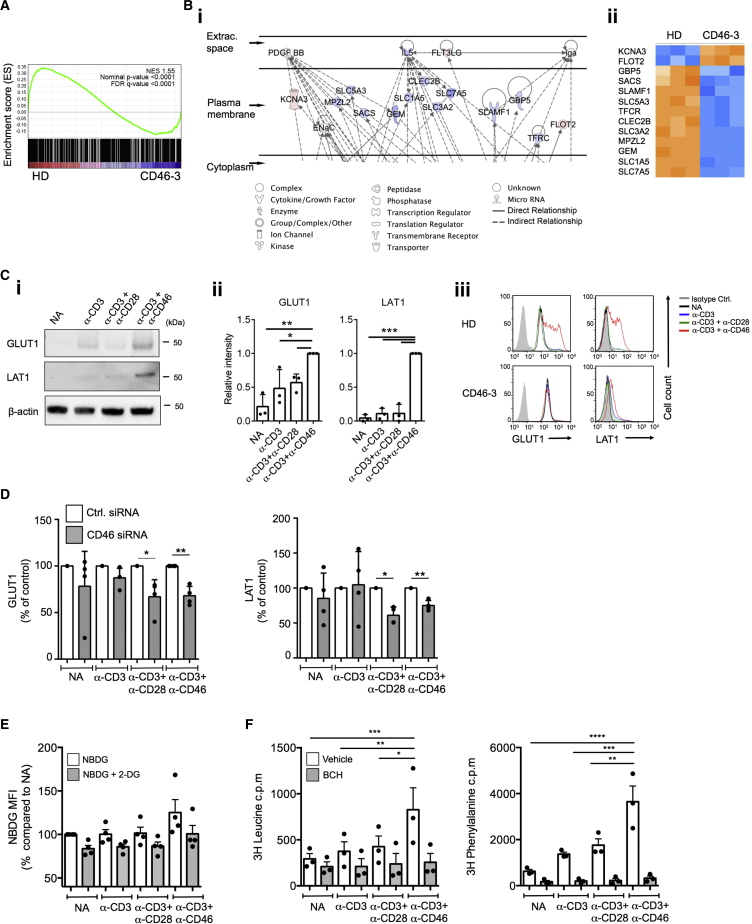
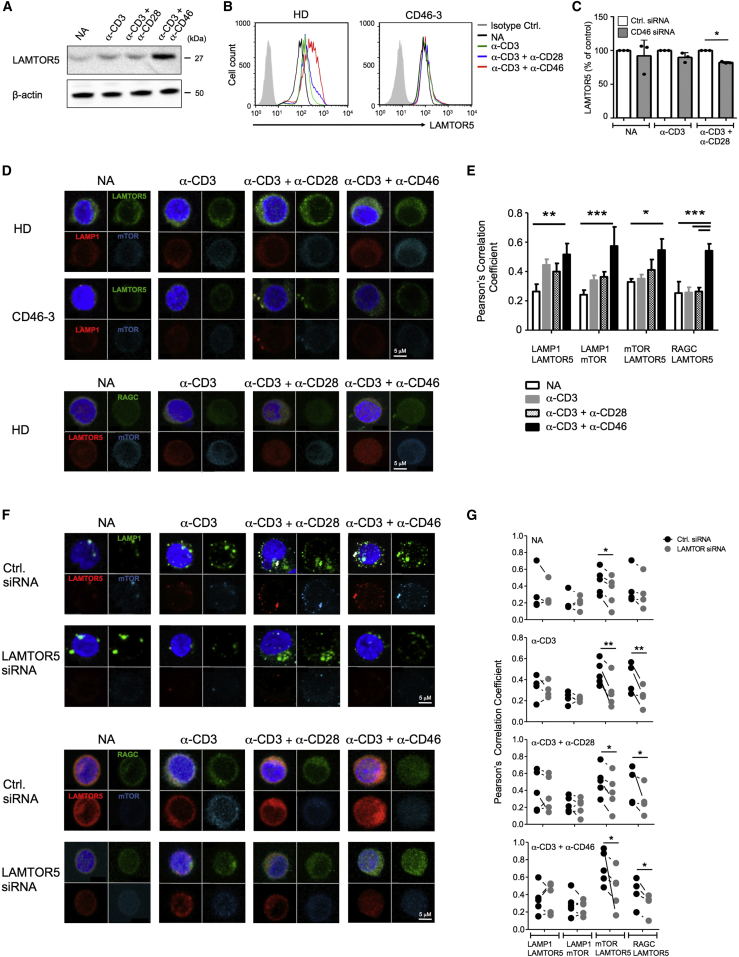
Expansion and acquisition of Th1 cell effector function requires metabolic reprogramming; however, the signals instructing these adaptations remain poorly defined. Here we found that in activated human T cells, autocrine stimulation of the complement receptor CD46, and specifically its intracellular domain CYT-1, was required for induction of the amino acid (AA) transporter LAT1 and enhanced expression of the glucose transporter GLUT1. Furthermore, CD46 activation simultaneously drove expression of LAMTOR5, which mediated assembly of the AA-sensing Ragulator-Rag-mTORC1 complex and increased glycolysis and oxidative phosphorylation (OXPHOS), required for cytokine production. T cells from CD46-deficient patients, characterized by defective Th1 cell induction, failed to upregulate the molecular components of this metabolic program as well as glycolysis and OXPHOS, but IFN-γ production could be reinstated by retrovirus-mediated CD46-CYT-1 expression. These data establish a critical link between the complement system and immunometabolic adaptations driving human CD4(+) T cell effector function.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures





Comment in
-
Complementing T Cells' Functions: Bringing in Metabolism Matters.Immunity. 2015 Jun 16;42(6):977-9. doi: 10.1016/j.immuni.2015.06.008. Immunity. 2015. PMID: 26084014
References
-
- Astier A., Trescol-Biémont M.C., Azocar O., Lamouille B., Rabourdin-Combe C. Cutting edge: CD46, a new costimulatory molecule for T cells, that induces p120CBL and LAT phosphorylation. J. Immunol. 2000;164:6091–6095. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
