

Florigen-Encoding Genes of Day-Neutral and Photoperiod-Sensitive Maize Are Regulated by Different Chromatin Modifications at the Floral Transition
- PMID: 26084920
- PMCID: PMC4528754
- DOI: 10.1104/pp.15.00535
Florigen-Encoding Genes of Day-Neutral and Photoperiod-Sensitive Maize Are Regulated by Different Chromatin Modifications at the Floral Transition
Abstract
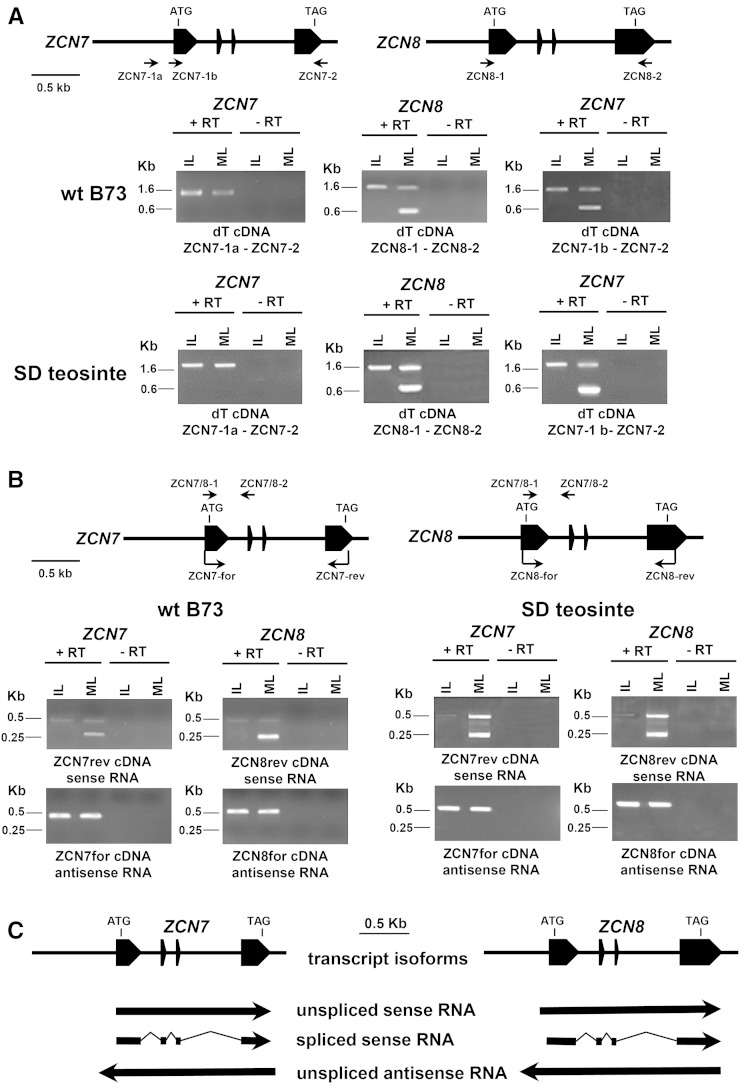
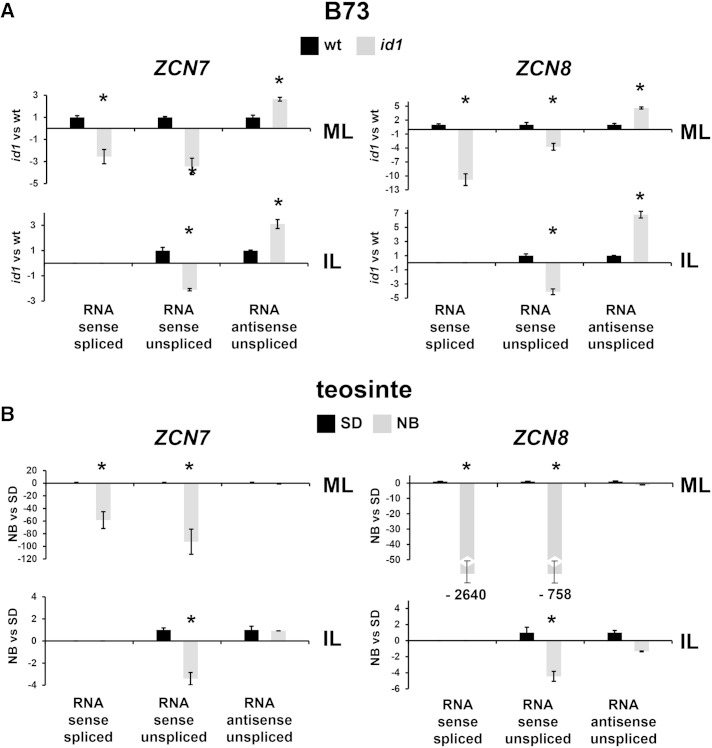
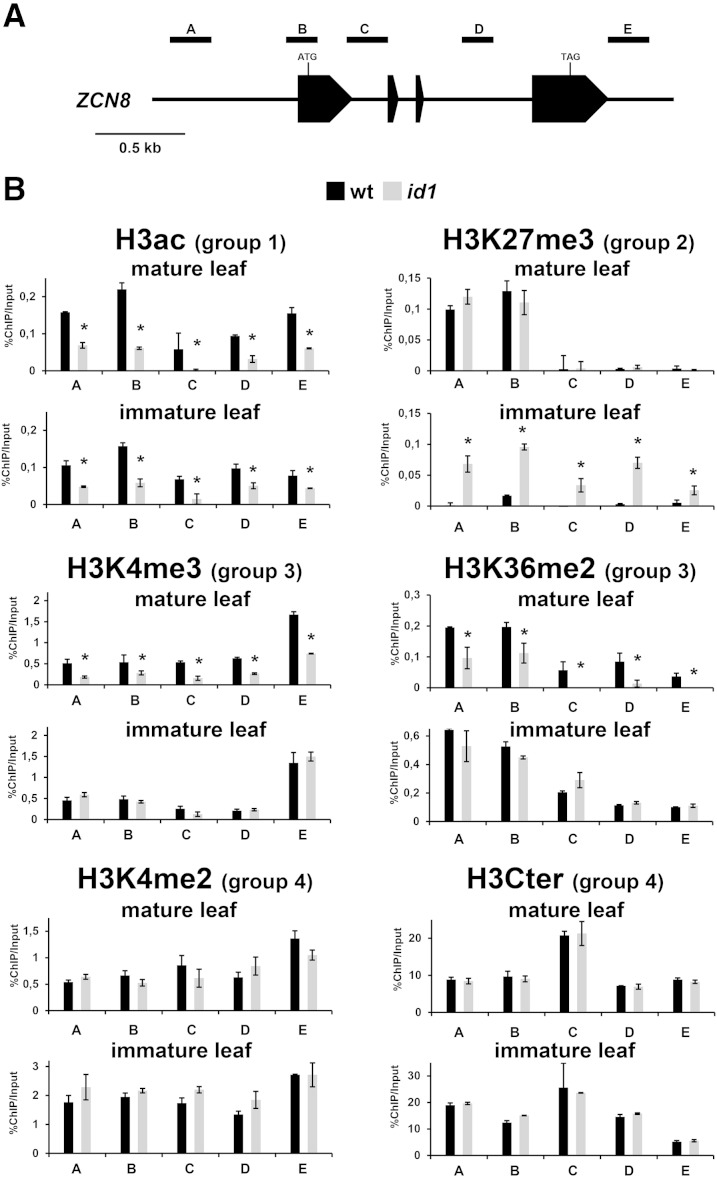
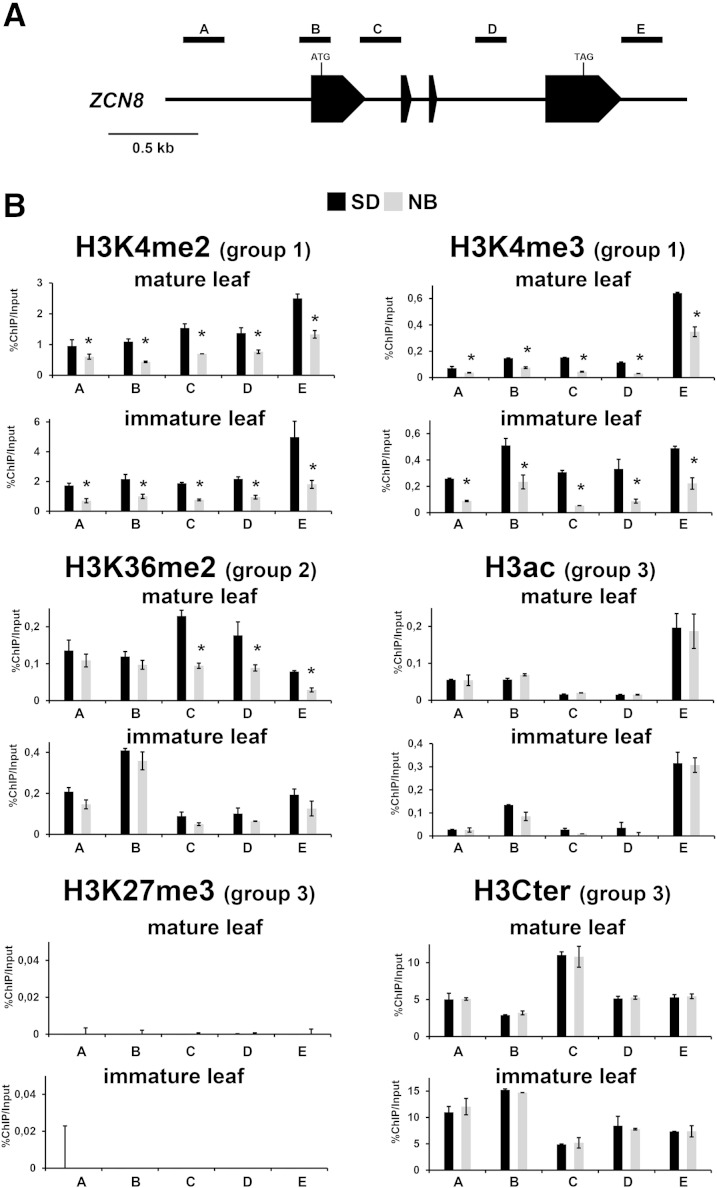
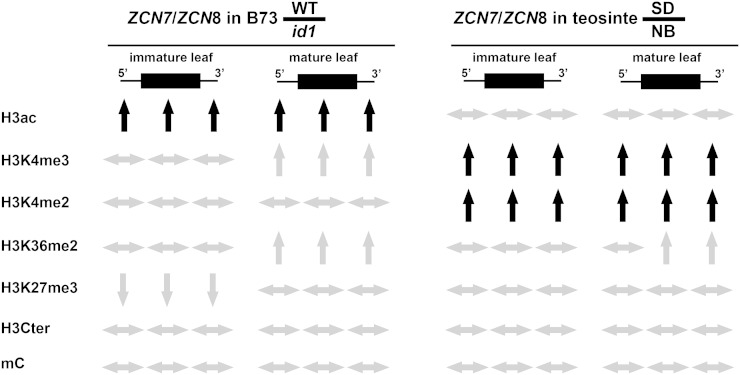
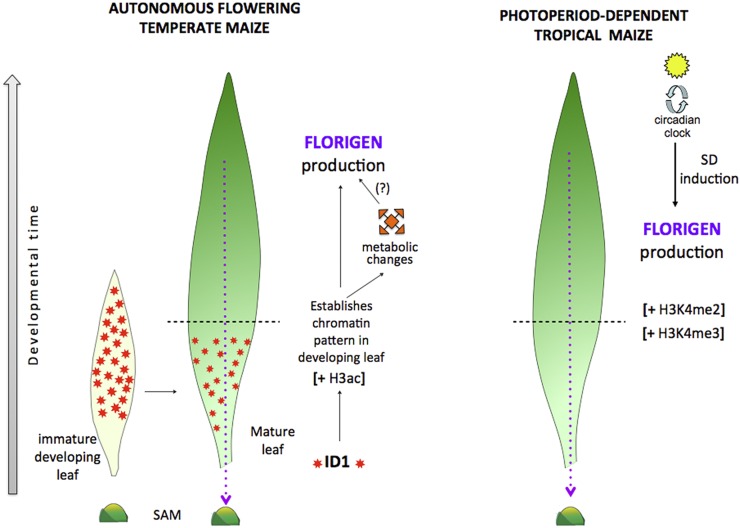
The activity of the maize (Zea mays) florigen gene ZEA CENTRORADIALIS8 (ZCN8) is associated with the floral transition in both day-neutral temperate maize and short-day (SD)-requiring tropical maize. We analyzed transcription and chromatin modifications at the ZCN8 locus and its nearly identical paralog ZCN7 during the floral transition. This analysis was performed with day-neutral maize (Zea mays ssp. mays), where flowering is promoted almost exclusively via the autonomous pathway through the activity of the regulatory gene indeterminate1 (id1), and tropical teosinte (Zea mays ssp. parviglumis) under floral inductive and noninductive photoperiods. Comparison of ZCN7/ZCN8 histone modification profiles in immature leaves of nonflowering id1 mutants and teosinte grown under floral inhibitory photoperiods reveals that both id1 floral inductive activity and SD-mediated induction result in histone modification patterns that are compatible with the formation of transcriptionally competent chromatin environments. Specific histone modifications are maintained during leaf development and may represent a chromatin signature that favors the production of processed ZCN7/ZCN8 messenger RNA in florigen-producing mature leaf. However, whereas id1 function promotes histone H3 hyperacetylation, SD induction is associated with increased histone H3 dimethylation and trimethylation at lysine-4. In addition, id1 and SD differently affect the production of ZCN7/ZCN8 antisense transcript. These observations suggest that distinct mechanisms distinguish florigen regulation in response to autonomous and photoperiod pathways. Finally, the identical expression and histone modification profiles of ZCN7 and ZCN8 in response to floral induction suggest that ZCN7 may represent a second maize florigen.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures

Similar articles
-
ZCN8 encodes a potential orthologue of Arabidopsis FT florigen that integrates both endogenous and photoperiod flowering signals in maize.J Exp Bot. 2011 Oct;62(14):4833-42. doi: 10.1093/jxb/err129. Epub 2011 Jul 5. J Exp Bot. 2011. PMID: 21730358 Free PMC article.
-
Transcript and metabolite signature of maize source leaves suggests a link between transitory starch to sucrose balance and the autonomous floral transition.J Exp Bot. 2012 Sep;63(14):5079-92. doi: 10.1093/jxb/ers158. Epub 2012 Jul 12. J Exp Bot. 2012. PMID: 22791826 Free PMC article.
-
Expression differences between normal and indeterminate1 maize suggest downstream targets of ID1, a floral transition regulator in maize.J Exp Bot. 2007;58(13):3679-93. doi: 10.1093/jxb/erm217. Epub 2007 Oct 10. J Exp Bot. 2007. PMID: 17928372
-
The coincidence of critical day length recognition for florigen gene expression and floral transition under long-day conditions in rice.Mol Plant. 2013 May;6(3):635-49. doi: 10.1093/mp/sst022. Epub 2013 Feb 15. Mol Plant. 2013. PMID: 23416454 Review.
-
Florigen in rice: complex gene network for florigen transcription, florigen activation complex, and multiple functions.Curr Opin Plant Biol. 2013 May;16(2):228-35. doi: 10.1016/j.pbi.2013.01.005. Epub 2013 Feb 28. Curr Opin Plant Biol. 2013. PMID: 23453779 Review.
Cited by
-
Coordinative regulation of plants growth and development by light and circadian clock.aBIOTECH. 2021 Mar 27;2(2):176-189. doi: 10.1007/s42994-021-00041-6. eCollection 2021 Jun. aBIOTECH. 2021. PMID: 36304756 Free PMC article. Review.
-
Understanding Past, and Predicting Future, Niche Transitions based on Grass Flowering Time Variation.Plant Physiol. 2020 Jul;183(3):822-839. doi: 10.1104/pp.20.00100. Epub 2020 May 13. Plant Physiol. 2020. PMID: 32404414 Free PMC article. Review.
-
INDETERMINATE1-mediated expression of FT family genes is required for proper timing of flowering in Brachypodium distachyon.Proc Natl Acad Sci U S A. 2023 Nov 14;120(46):e2312052120. doi: 10.1073/pnas.2312052120. Epub 2023 Nov 7. Proc Natl Acad Sci U S A. 2023. PMID: 37934817 Free PMC article.
-
Major niche transitions in Pooideae correlate with variation in photoperiodic flowering and evolution of CCT domain genes.J Exp Bot. 2022 Jun 24;73(12):4079-4093. doi: 10.1093/jxb/erac149. J Exp Bot. 2022. PMID: 35394528 Free PMC article.
-
Epigenetics and its role in effecting agronomical traits.Front Plant Sci. 2022 Aug 15;13:925688. doi: 10.3389/fpls.2022.925688. eCollection 2022. Front Plant Sci. 2022. PMID: 36046583 Free PMC article. Review.
References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309: 1052–1056 - PubMed
-
- Andrés F, Coupland G (2012) The genetic basis of flowering responses to seasonal cues. Nat Rev Genet 13: 627–639 - PubMed
-
- Bouchet S, Servin B, Bertin P, Madur D, Combes V, Dumas F, Brunel D, Laborde J, Charcosset A, Nicolas S (2013) Adaptation of maize to temperate climates: mid-density genome-wide association genetics and diversity patterns reveal key genomic regions, with a major contribution of the Vgt2 (ZCN8) locus. PLoS ONE 8: e71377. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
