

Porphyromonas gingivalis Stimulates Bone Resorption by Enhancing RANKL (Receptor Activator of NF-κB Ligand) through Activation of Toll-like Receptor 2 in Osteoblasts
- PMID: 26085099
- PMCID: PMC4536425
- DOI: 10.1074/jbc.M115.655787
Porphyromonas gingivalis Stimulates Bone Resorption by Enhancing RANKL (Receptor Activator of NF-κB Ligand) through Activation of Toll-like Receptor 2 in Osteoblasts
Abstract
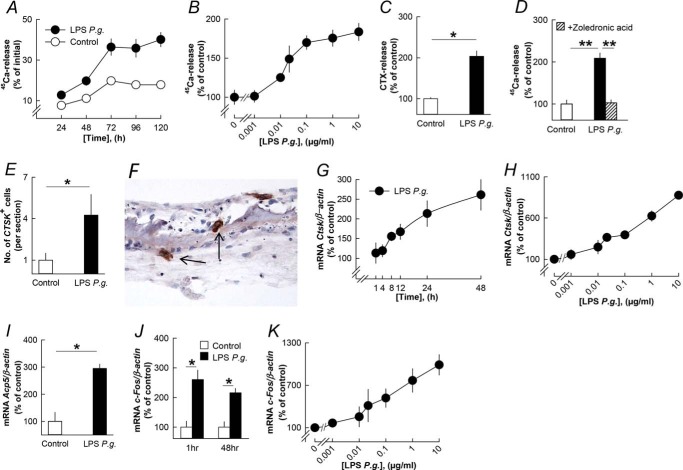
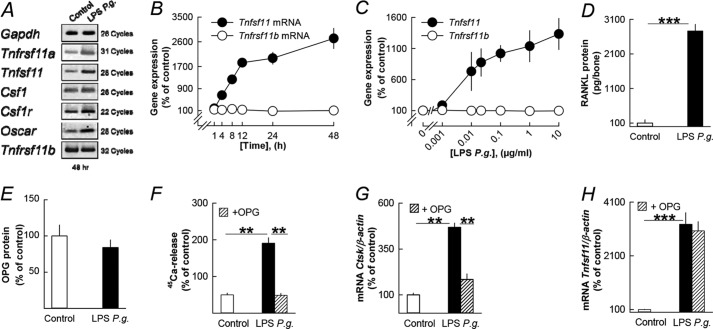
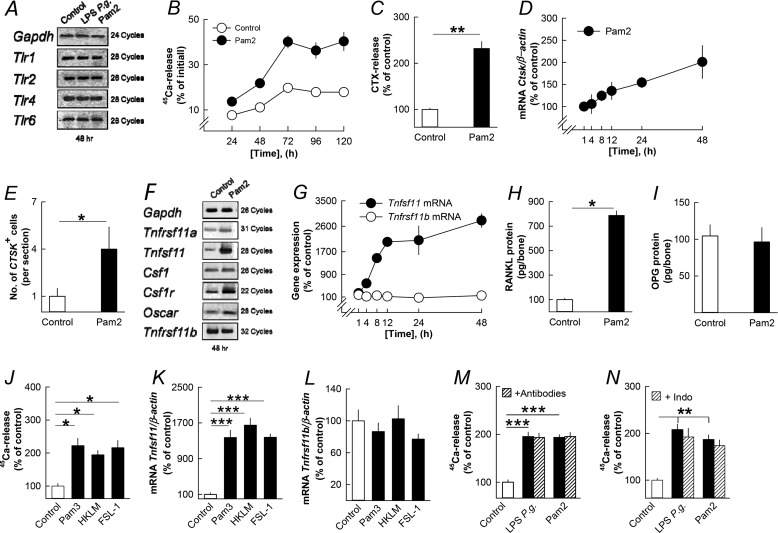
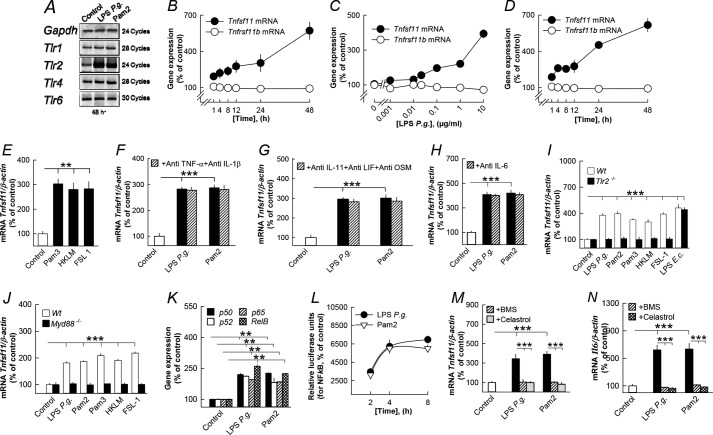
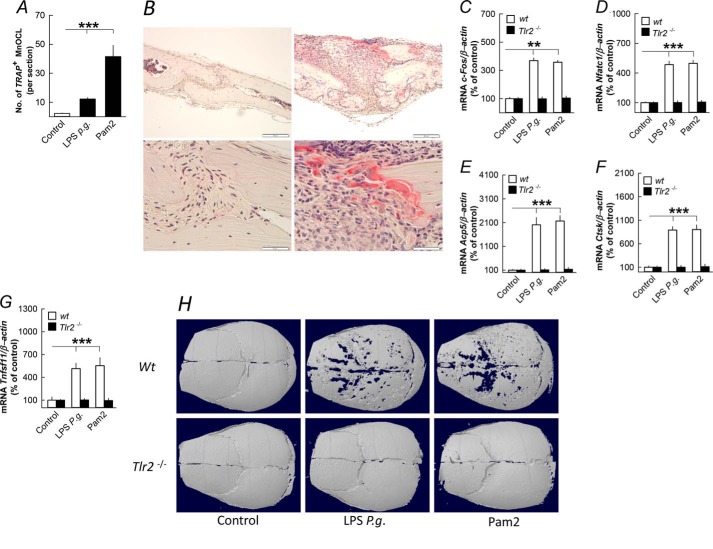
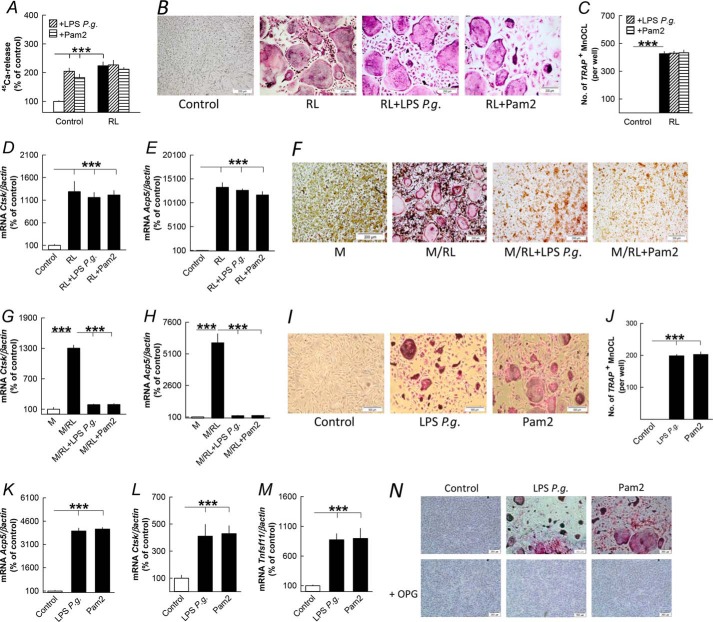
Periodontitis has been associated with rheumatoid arthritis. In experimental arthritis, concomitant periodontitis caused by oral infection with Porphyromonas gingivalis enhances articular bone loss. The aim of this study was to investigate how lipopolysaccharide (LPS) from P. gingivalis stimulates bone resorption. The effects by LPS P. gingivalis and four other TLR2 ligands on bone resorption, osteoclast formation, and gene expression in wild type and Tlr2-deficient mice were assessed in ex vivo cultures of mouse parietal bones and in an in vivo model in which TLR2 agonists were injected subcutaneously over the skull bones. LPS P. gingivalis stimulated mineral release and matrix degradation in the parietal bone organ cultures by increasing differentiation and formation of mature osteoclasts, a response dependent on increased RANKL (receptor activator of NF-κB ligand). LPS P. gingivalis stimulated RANKL in parietal osteoblasts dependent on the presence of TLR2 and through a MyD88 and NF-κB-mediated mechanism. Similarly, the TLR2 agonists HKLM, FSL1, Pam2, and Pam3 stimulated RANKL in osteoblasts and parietal bone resorption. LPS P. gingivalis and Pam2 robustly enhanced osteoclast formation in periosteal/endosteal cell cultures by increasing RANKL. LPS P. gingivalis and Pam2 also up-regulated RANKL and osteoclastic genes in vivo, resulting in an increased number of periosteal osteoclasts and immense bone loss in wild type mice but not in Tlr2-deficient mice. These data demonstrate that LPS P. gingivalis stimulates periosteal osteoclast formation and bone resorption by stimulating RANKL in osteoblasts via TLR2. This effect might be important for periodontal bone loss and for the enhanced bone loss seen in rheumatoid arthritis patients with concomitant periodontal disease.
Keywords: bone; inflammation; innate immunity; osteoblast; osteoclast; toll-like receptor (TLR).
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Walsh N. C., Gravallese E. M. (2010) Bone remodeling in rheumatic disease: a question of balance. Immunol. Rev. 233, 301–312 - PubMed
-
- Bartold P. M., Cantley M. D., Haynes D. R. (2010) Mechanisms and control of pathologic bone loss in periodontitis. Periodontol. 2000 53, 55–69 - PubMed
-
- Souza P. P., Lerner U. H. (2013) The role of cytokines in inflammatory bone loss. Immunol Invest. 42, 555–622 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
