

Mia40 Protein Serves as an Electron Sink in the Mia40-Erv1 Import Pathway
- PMID: 26085103
- PMCID: PMC4543643
- DOI: 10.1074/jbc.M115.669440
Mia40 Protein Serves as an Electron Sink in the Mia40-Erv1 Import Pathway
Abstract
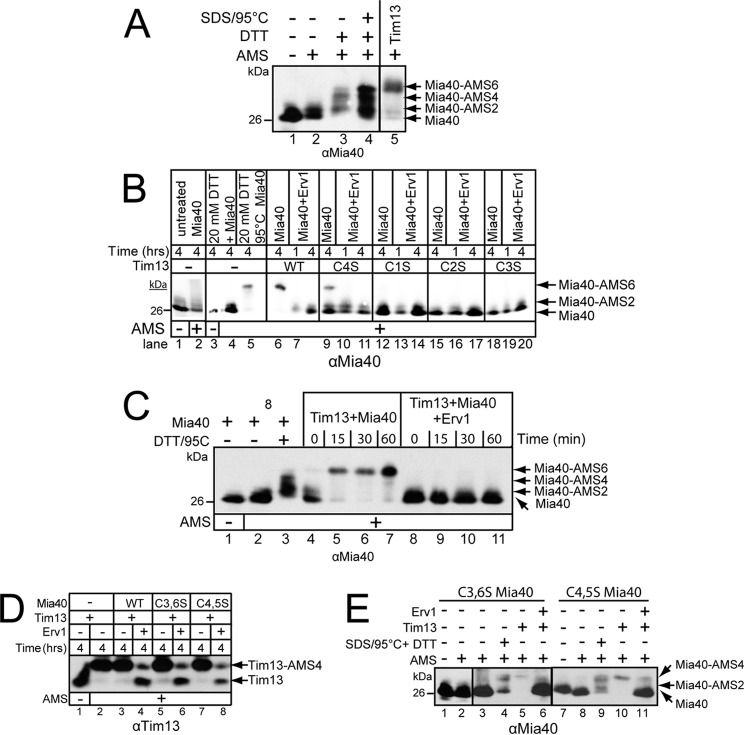
A redox-regulated import pathway consisting of Mia40 and Erv1 mediates the import of cysteine-rich proteins into the mitochondrial intermembrane space. Mia40 is the oxidoreductase that inserts two disulfide bonds into the substrate simultaneously. However, Mia40 has one redox-active cysteine pair, resulting in ambiguity about how Mia40 accepts numerous electrons during substrate oxidation. In this study, we have addressed the oxidation of Tim13 in vitro and in organello. Reductants such as glutathione and ascorbate inhibited both the oxidation of the substrate Tim13 in vitro and the import of Tim13 and Cmc1 into isolated mitochondria. In addition, a ternary complex consisting of Erv1, Mia40, and substrate, linked by disulfide bonds, was not detected in vitro. Instead, Mia40 accepted six electrons from substrates, and this fully reduced Mia40 was sensitive to protease, indicative of conformational changes in the structure. Mia40 in mitochondria from the erv1-101 mutant was also trapped in a completely reduced state, demonstrating that Mia40 can accept up to six electrons as substrates are imported. Therefore, these studies support that Mia40 functions as an electron sink to facilitate the insertion of two disulfide bonds into substrates.
Keywords: disulfide; mitochondria; oxidation-reduction (redox); protein import; redox; redox regulation; thiol.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






References
-
- Stojanovski D., Bragoszewski P., Chacinska A. (2012) The MIA pathway: a tight bond between protein transport and oxidative folding in mitochondria. Biochim. Biophys. Acta 1823, 1142–1150 - PubMed
-
- Koehler C. M., Merchant S., Schatz G. (1999) How membrane proteins travel across the mitochondrial intermembrane space. Trends Biochem. Sci. 24, 428–432 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
