

Assembly of Epstein-Barr Virus Capsid in Promyelocytic Leukemia Nuclear Bodies
- PMID: 26085145
- PMCID: PMC4524052
- DOI: 10.1128/JVI.01114-15
Assembly of Epstein-Barr Virus Capsid in Promyelocytic Leukemia Nuclear Bodies
Abstract
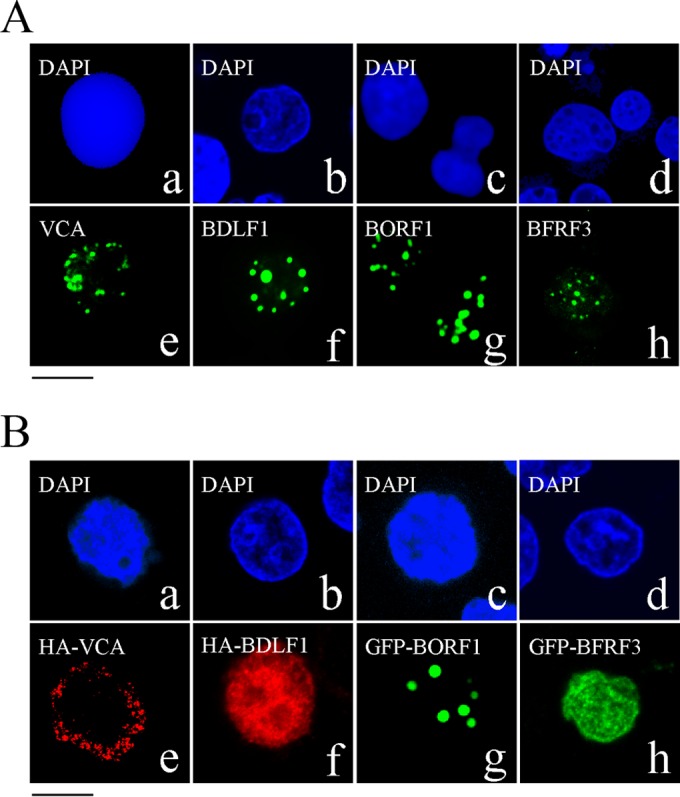
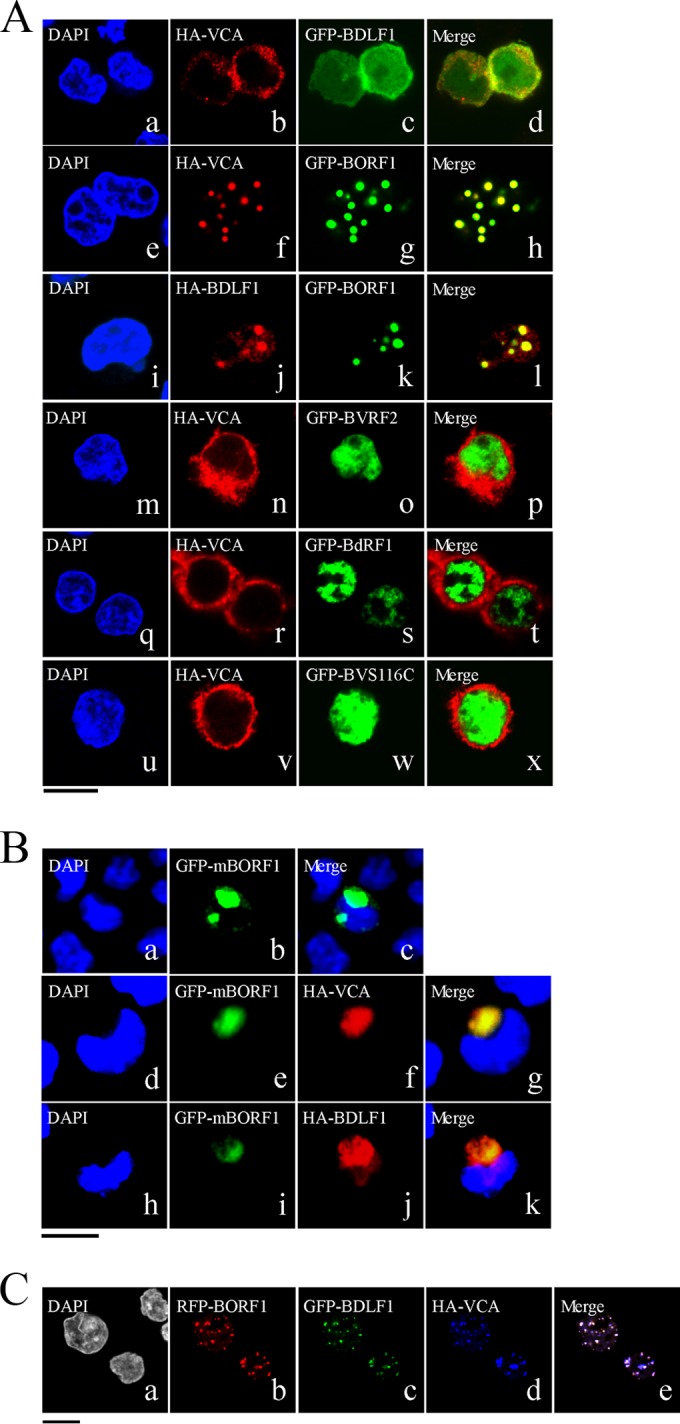
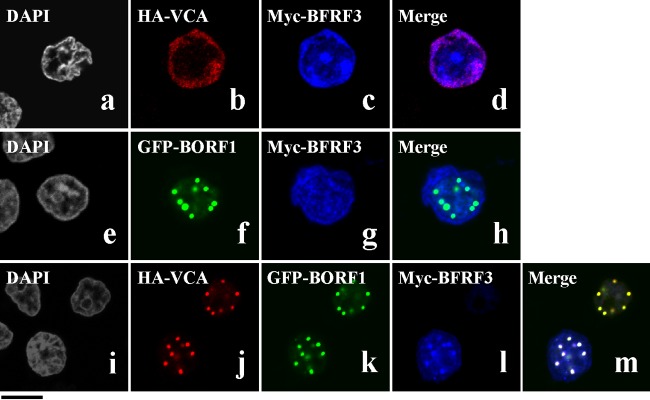
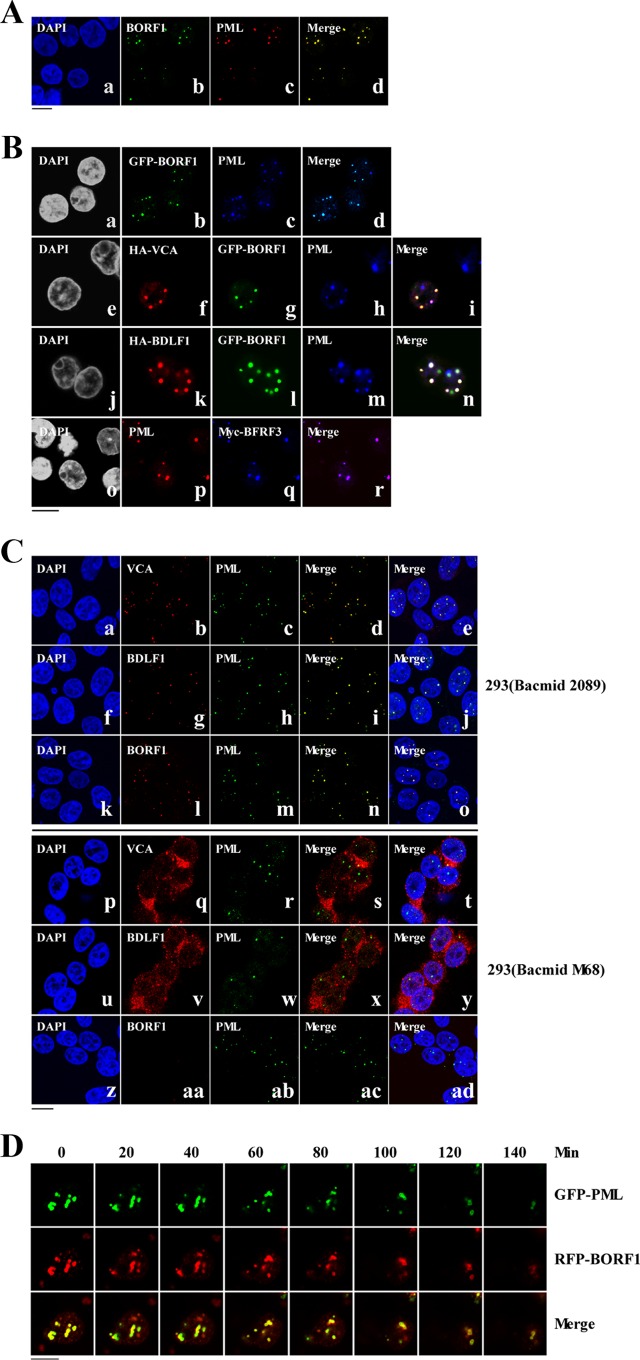
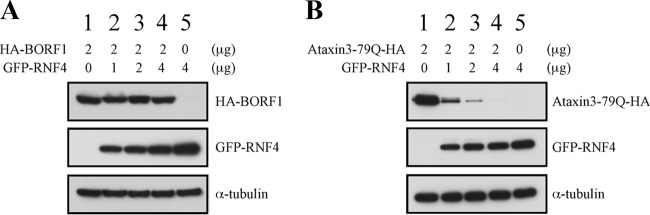
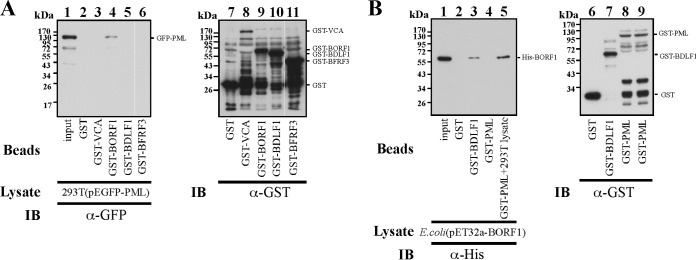
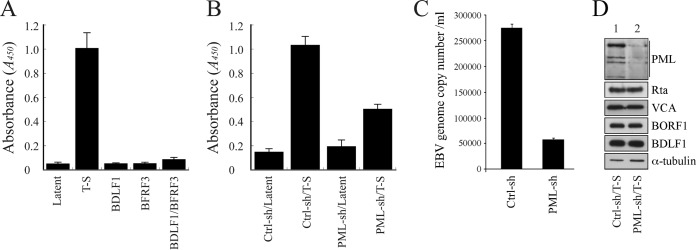
The Epstein-Barr virus (EBV) capsid contains a major capsid protein, VCA; two minor capsid proteins, BDLF1 and BORF1; and a small capsid protein, BFRF3. During the lytic cycle, these capsid proteins are synthesized and imported into the host nucleus for capsid assembly. This study finds that EBV capsid proteins colocalize with promyelocytic leukemia (PML) nuclear bodies (NBs) in P3HR1 cells during the viral lytic cycle, appearing as nuclear speckles under a confocal laser scanning microscope. In a glutathione S-transferase pulldown study, we show that BORF1 interacts with PML-NBs in vitro. BORF1 also colocalizes with PML-NBs in EBV-negative Akata cells after transfection and is responsible for bringing VCA and the VCA-BFRF3 complex from the cytoplasm to PML-NBs in the nucleus. Furthermore, BDLF1 is dispersed throughout the cell when expressed alone but colocalizes with PML-NBs when BORF1 is also present in the cell. In addition, this study finds that knockdown of PML expression by short hairpin RNA does not influence the intracellular levels of capsid proteins but reduces the number of viral particles produced by P3HR1 cells. Together, these results demonstrate that BORF1 plays a critical role in bringing capsid proteins to PML-NBs, which may likely be the assembly sites of EBV capsids. The mechanisms elucidated in this study are critical to understanding the process of EBV capsid assembly. IMPORTANCE Capsid assembly is an important event during the Epstein-Barr virus (EBV) lytic cycle, as this process is required for the production of virions. In this study, confocal microscopy revealed that the EBV capsid protein BORF1 interacts with promyelocytic leukemia (PML) nuclear bodies (NBs) in the host nucleus and is responsible for transporting the other EBV capsid proteins, including VCA, BDLF1, and BFRF3, to these subnuclear locations prior to initiation of capsid assembly. This study also found that knockdown of PML expression by short hairpin RNA significantly reduces EBV capsid assembly capabilities. This enhanced understanding of capsid assembly offers potential for the development of novel antiviral strategies and therapies that can prevent the propagation and spread of EBV.
Figures
References
-
- Newcomb WW, Trus BL, Booy FP, Steven AC, Wall JS, Brown JC. 1993. Structure of the herpes simplex virus capsid. Molecular composition of the pentons and the triplexes. J Mol Biol 232:499–511. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
