

Blood Groups in Infection and Host Susceptibility
- PMID: 26085552
- PMCID: PMC4475644
- DOI: 10.1128/CMR.00109-14
Blood Groups in Infection and Host Susceptibility
Abstract
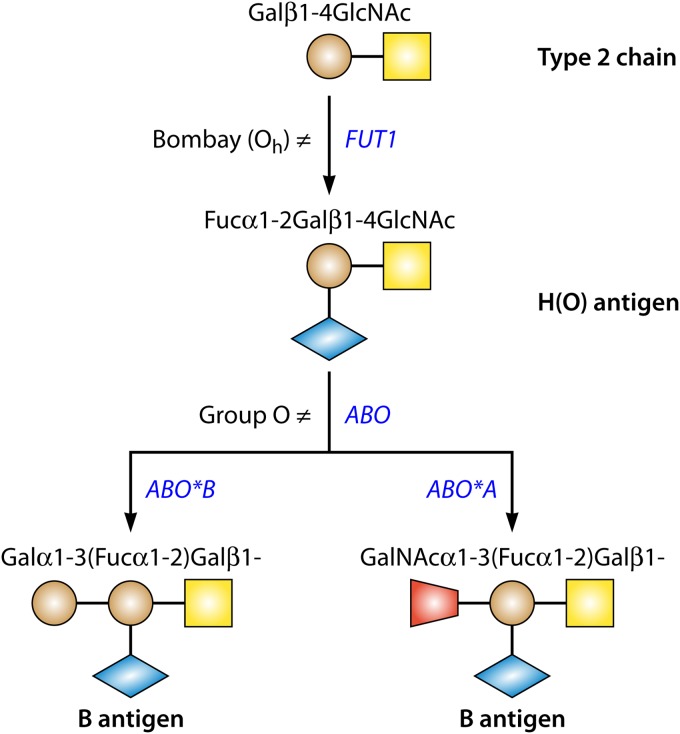
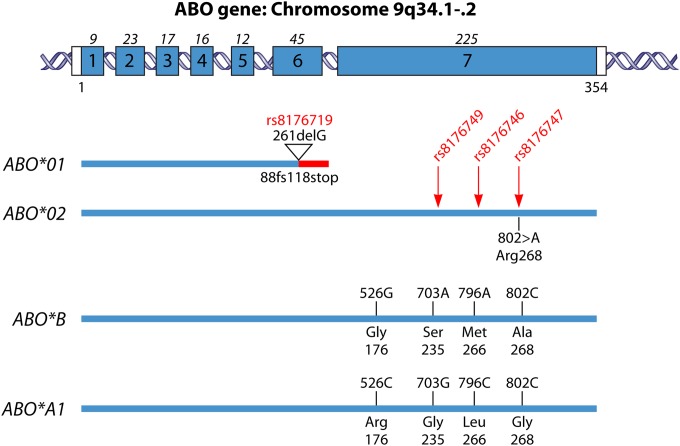
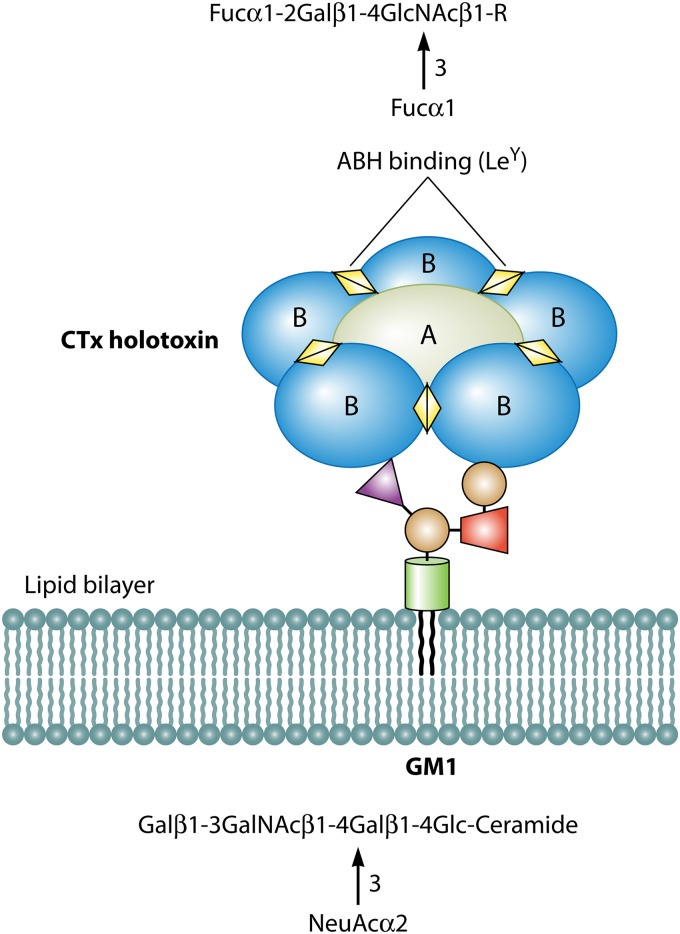
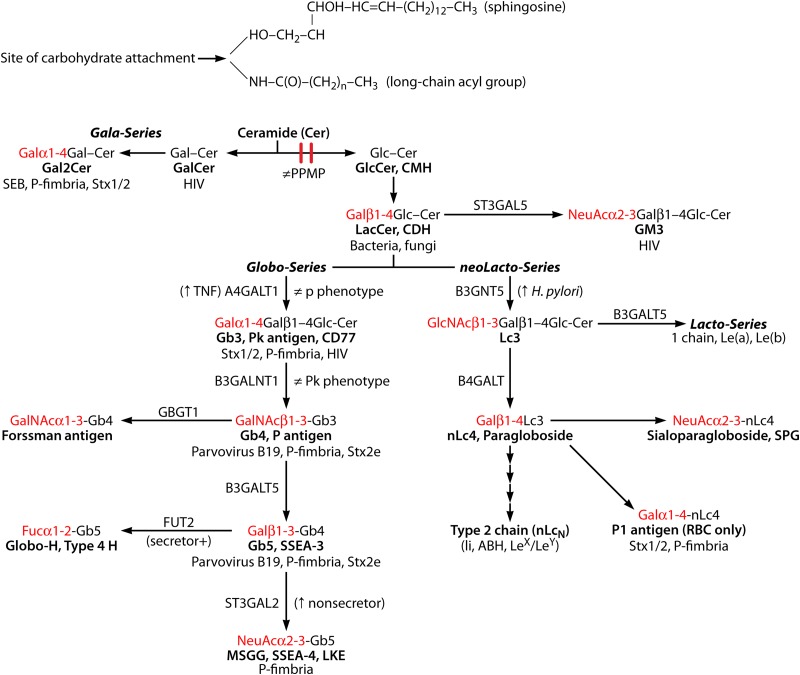
Blood group antigens represent polymorphic traits inherited among individuals and populations. At present, there are 34 recognized human blood groups and hundreds of individual blood group antigens and alleles. Differences in blood group antigen expression can increase or decrease host susceptibility to many infections. Blood groups can play a direct role in infection by serving as receptors and/or coreceptors for microorganisms, parasites, and viruses. In addition, many blood group antigens facilitate intracellular uptake, signal transduction, or adhesion through the organization of membrane microdomains. Several blood groups can modify the innate immune response to infection. Several distinct phenotypes associated with increased host resistance to malaria are overrepresented in populations living in areas where malaria is endemic, as a result of evolutionary pressures. Microorganisms can also stimulate antibodies against blood group antigens, including ABO, T, and Kell. Finally, there is a symbiotic relationship between blood group expression and maturation of the gastrointestinal microbiome.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

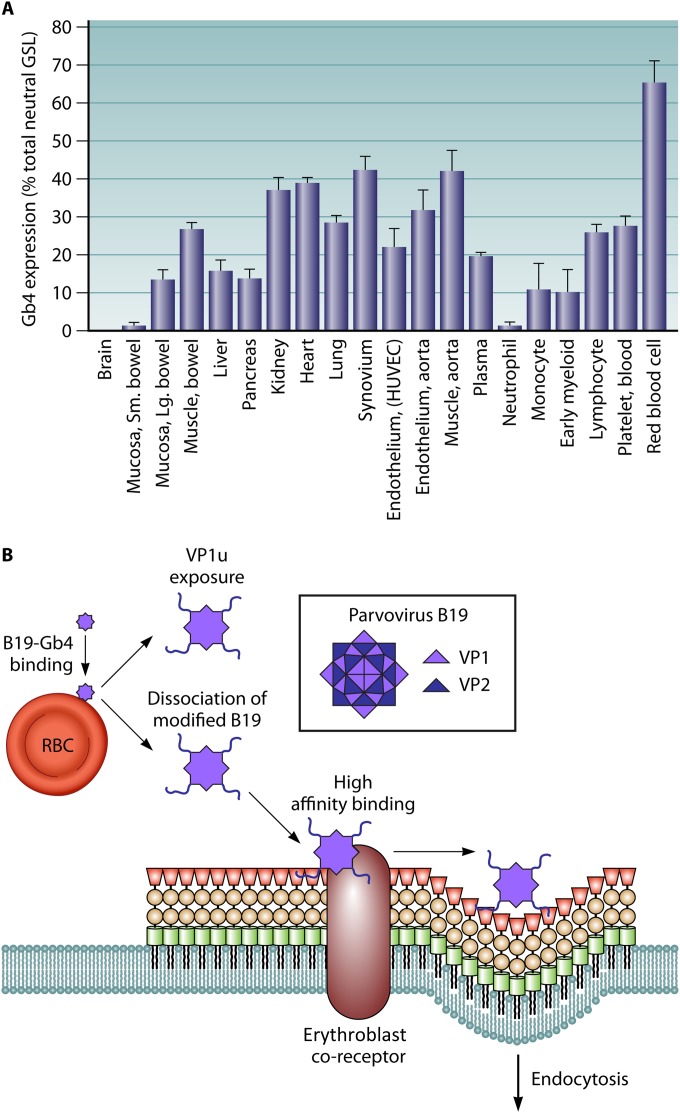
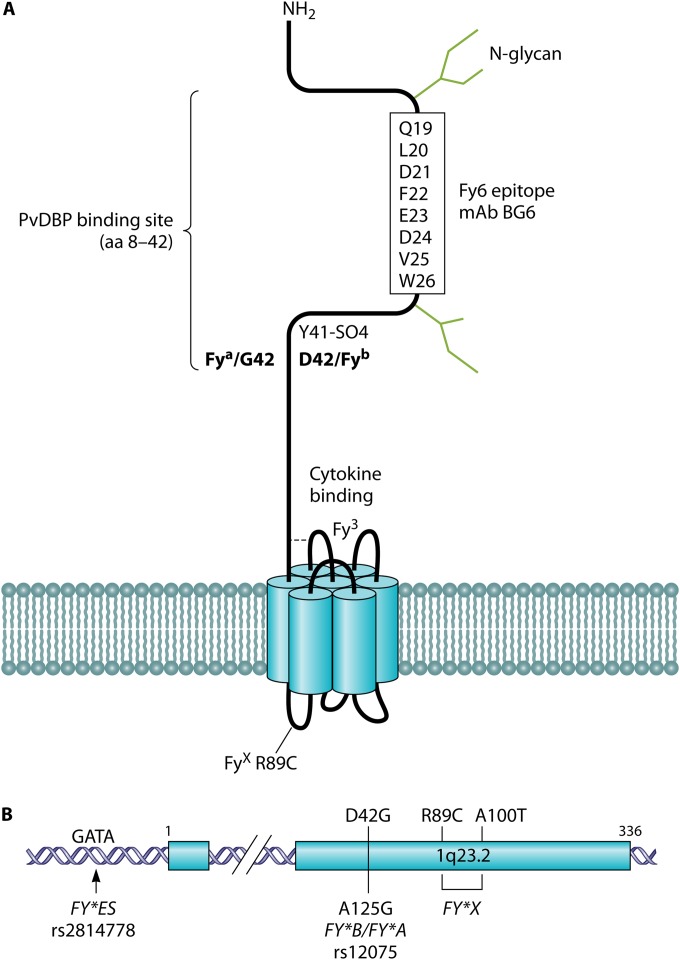
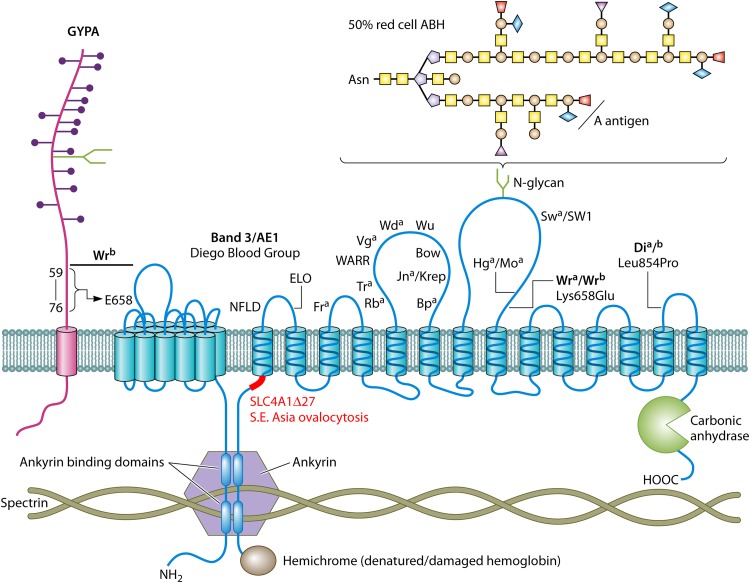
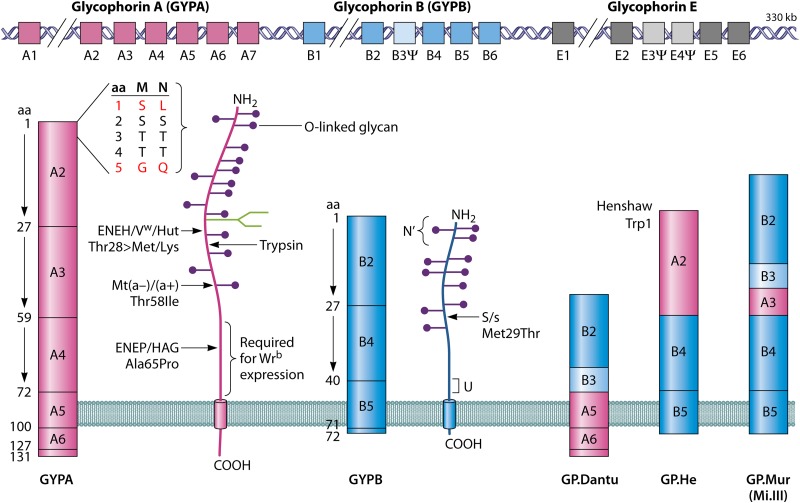
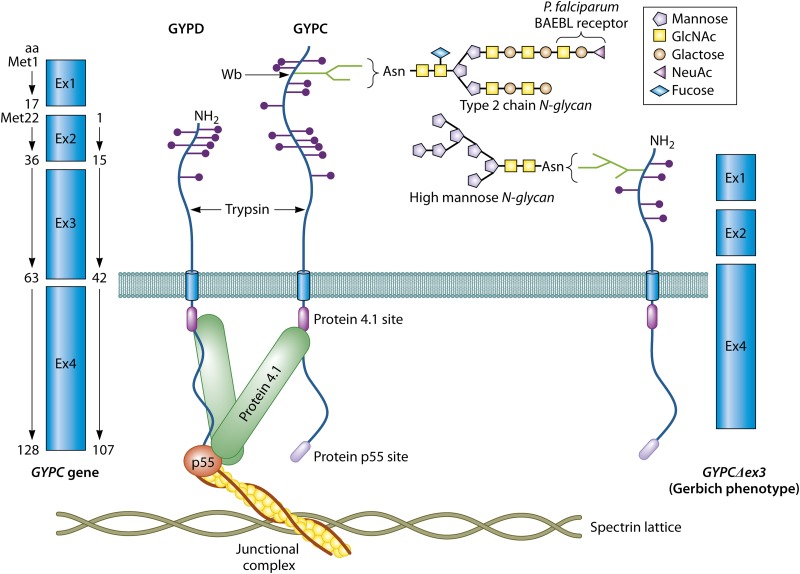
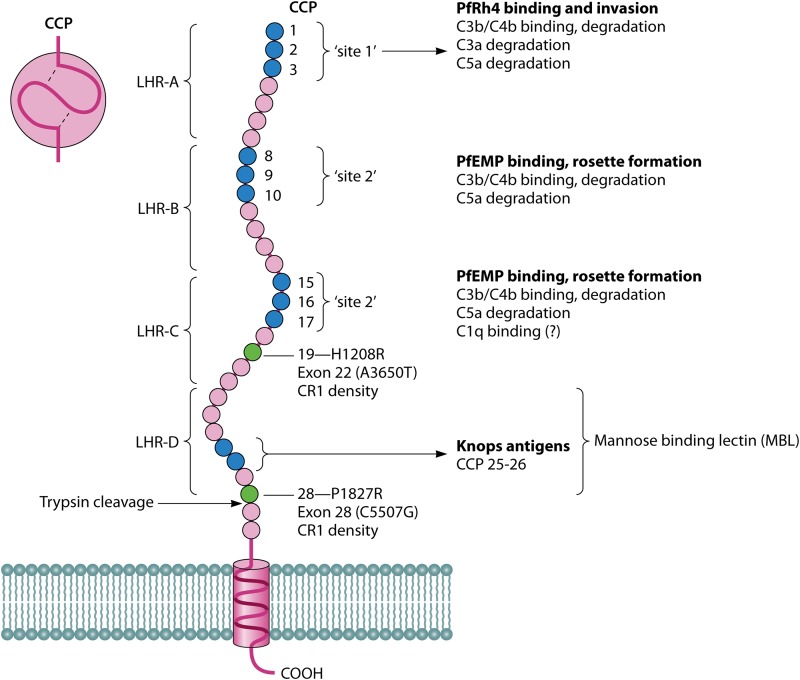
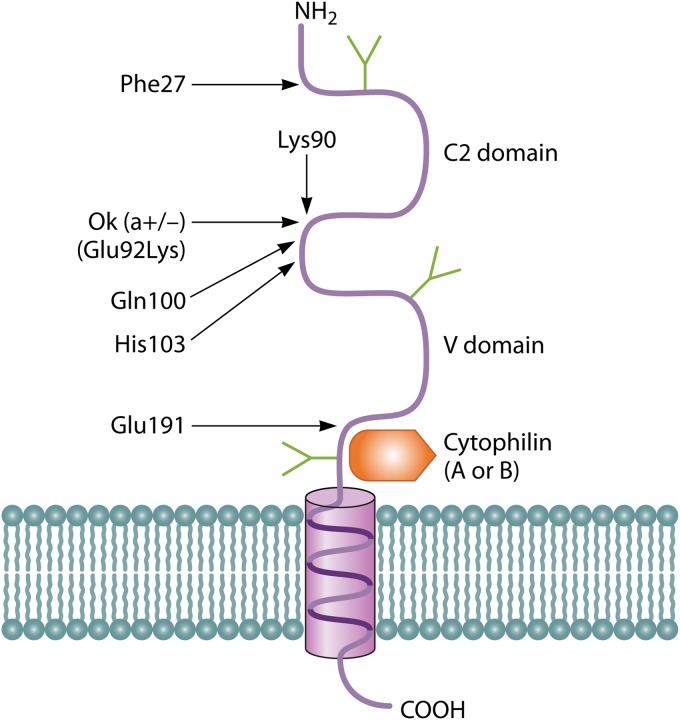
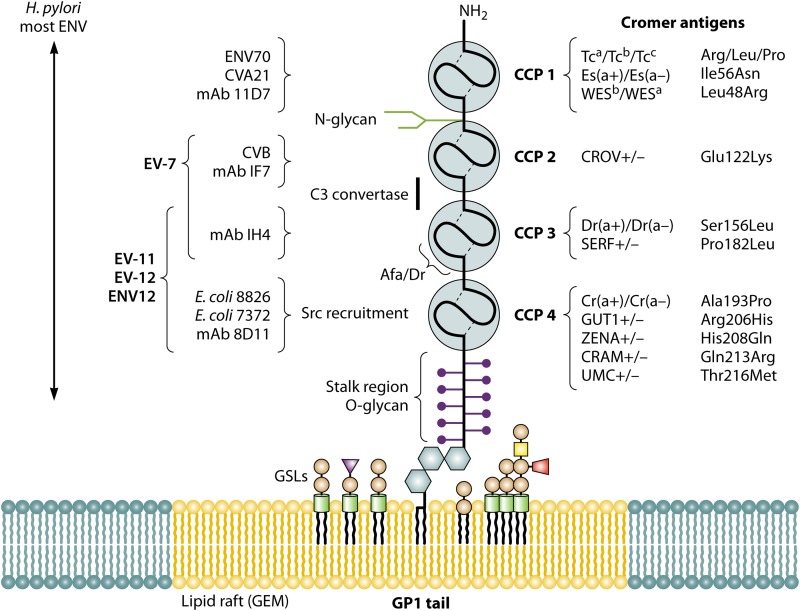
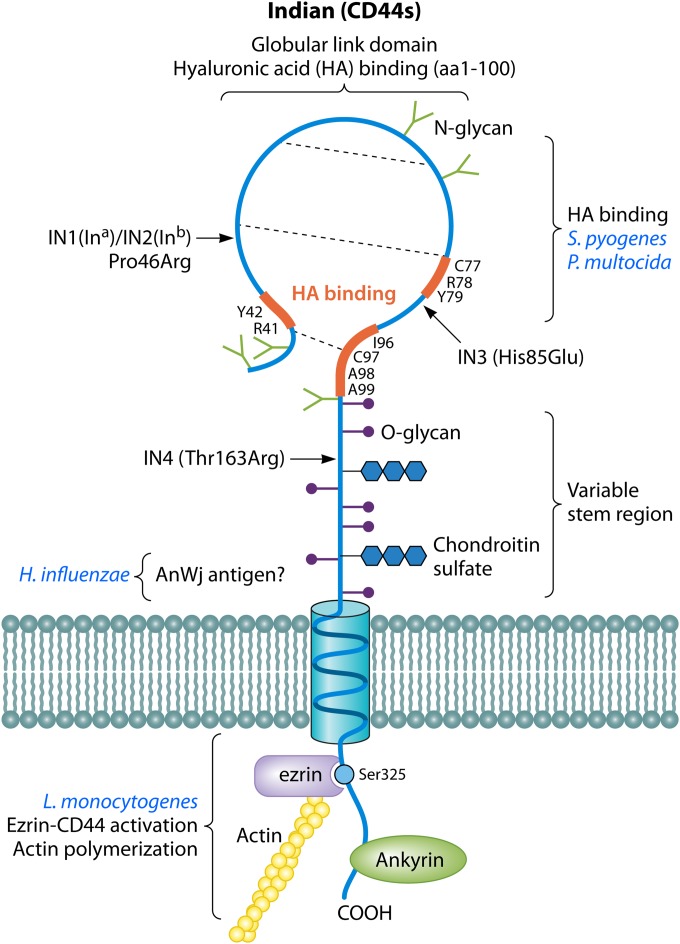
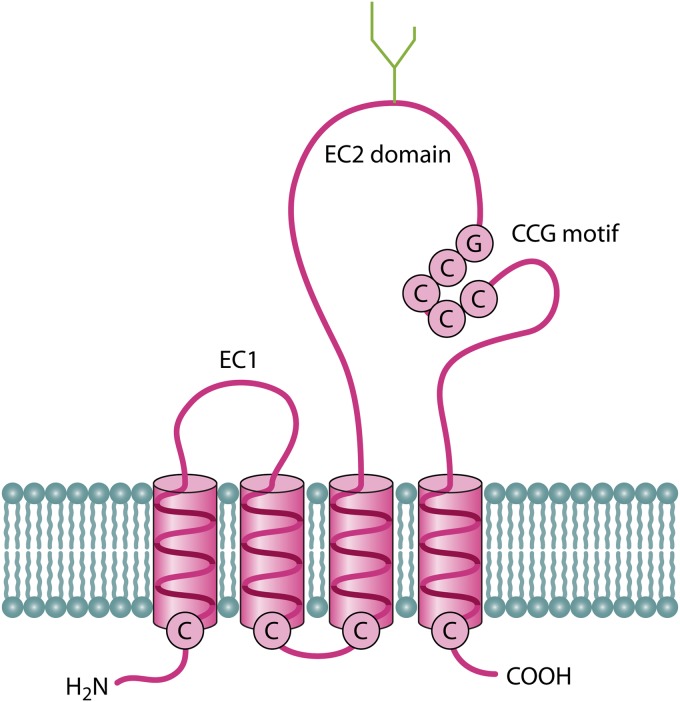

References
-
- Fung MK, Grossman BJ, Hillyer CD, Westhoff CM. 2014. Technical manual, 18th ed AABB Press, Bethesda, MD.
-
- Reid ME, Lomas-Francis C, Olsson ML. 2012. Blood group antigen FactsBook, 3rd ed Academic Press, Waltham, MA.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
