

Clonal Evolution and Blast Crisis Correlate with Enhanced Proteolytic Activity of Separase in BCR-ABL b3a2 Fusion Type CML under Imatinib Therapy
- PMID: 26087013
- PMCID: PMC4472749
- DOI: 10.1371/journal.pone.0129648
Clonal Evolution and Blast Crisis Correlate with Enhanced Proteolytic Activity of Separase in BCR-ABL b3a2 Fusion Type CML under Imatinib Therapy
Abstract
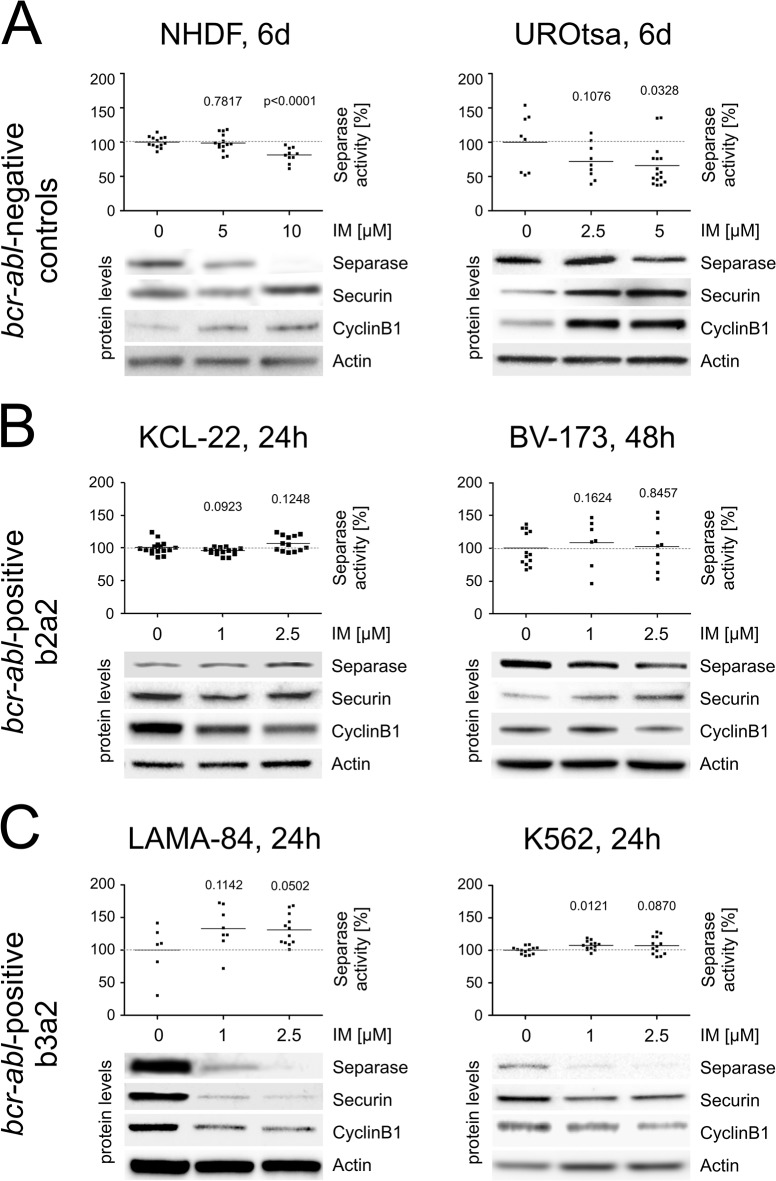
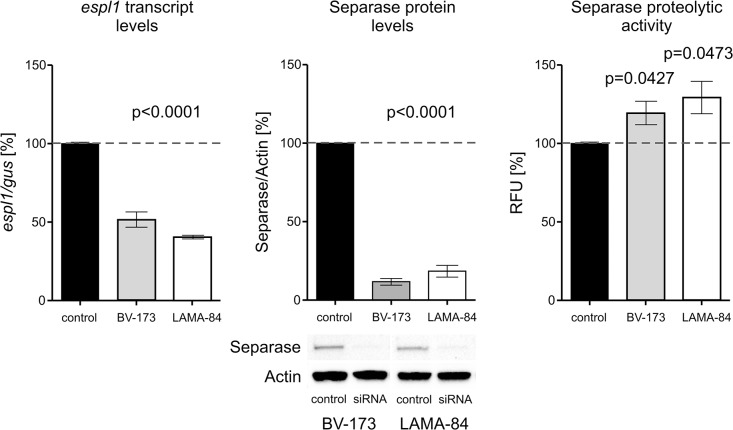
Unbalanced (major route) additional cytogenetic aberrations (ACA) at diagnosis of chronic myeloid leukemia (CML) indicate an increased risk of progression and shorter survival. Moreover, newly arising ACA under imatinib treatment and clonal evolution are considered features of acceleration and define failure of therapy according to the European LeukemiaNet (ELN) recommendations. On the basis of 1151 Philadelphia chromosome positive chronic phase patients of the randomized CML-study IV, we examined the incidence of newly arising ACA under imatinib treatment with regard to the p210BCR-ABL breakpoint variants b2a2 and b3a2. We found a preferential acquisition of unbalanced ACA in patients with b3a2 vs. b2a2 fusion type (ratio: 6.3 vs. 1.6, p = 0.0246) concurring with a faster progress to blast crisis for b3a2 patients (p = 0.0124). ESPL1/Separase, a cysteine endopeptidase, is a key player in chromosomal segregation during mitosis. Separase overexpression and/or hyperactivity has been reported from a wide range of cancers and cause defective mitotic spindles, chromosome missegregation and aneuploidy. We investigated the influence of p210BCR-ABL breakpoint variants and imatinib treatment on expression and proteolytic activity of Separase as measured with a specific fluorogenic assay on CML cell lines (b2a2: KCL-22, BV-173; b3a2: K562, LAMA-84). Despite a drop in Separase protein levels an up to 5.4-fold increase of Separase activity under imatinib treatment was observed exclusively in b3a2 but not in b2a2 cell lines. Mimicking the influence of imatinib on BV-173 and LAMA-84 cells by ESPL1 silencing stimulated Separase proteolytic activity in both b3a2 and b2a2 cell lines. Our data suggest the existence of a fusion type-related feedback mechanism that posttranslationally stimulates Separase proteolytic activity after therapy-induced decreases in Separase protein levels. This could render b3a2 CML cells more prone to aneuploidy and clonal evolution than b2a2 progenitors and may therefore explain the cytogenetic results of CML patients.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
