

SIGNAL TRANSDUCTION. Structural basis for nucleotide exchange in heterotrimeric G proteins
- PMID: 26089515
- PMCID: PMC4968074
- DOI: 10.1126/science.aaa5264
SIGNAL TRANSDUCTION. Structural basis for nucleotide exchange in heterotrimeric G proteins
Abstract
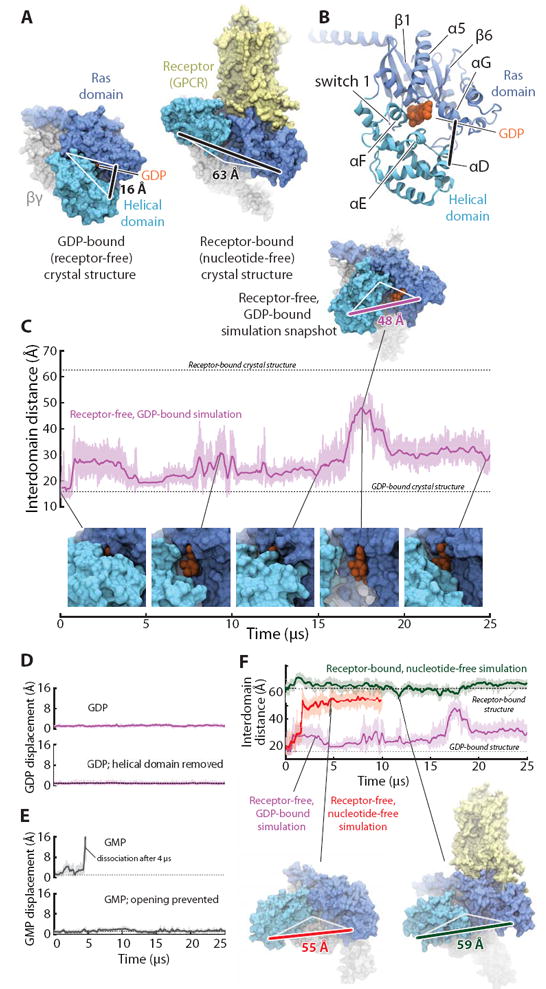
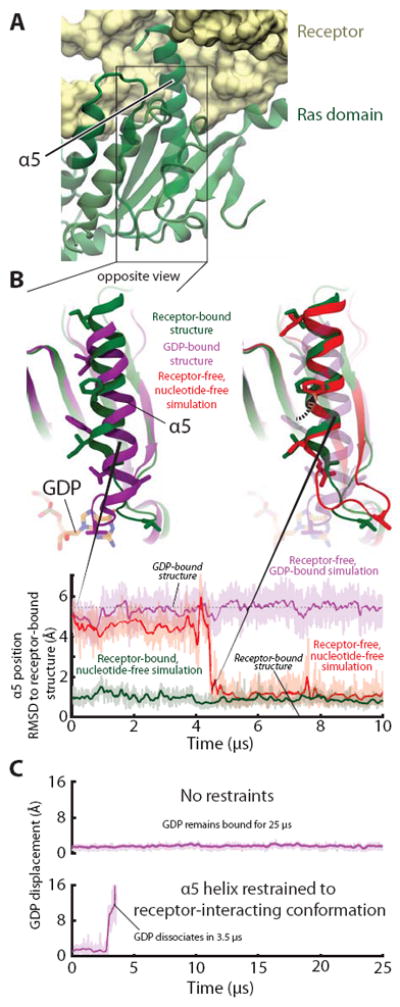
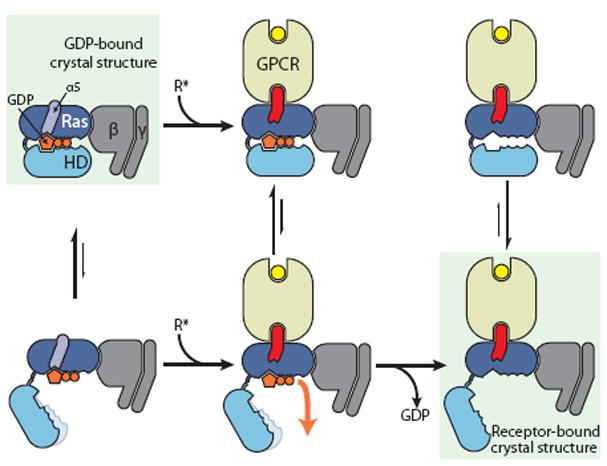
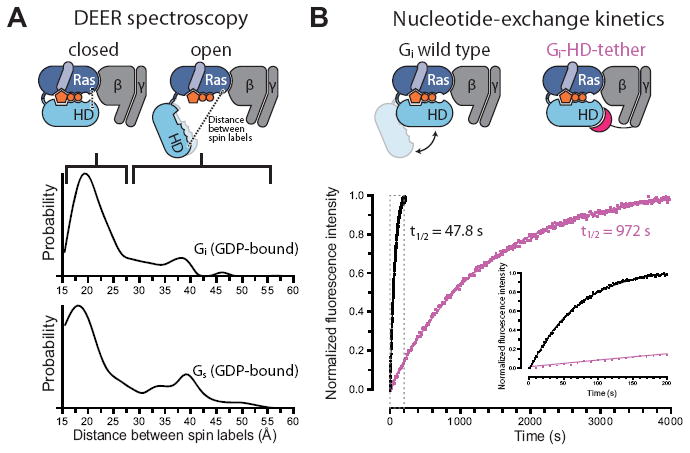
G protein-coupled receptors (GPCRs) relay diverse extracellular signals into cells by catalyzing nucleotide release from heterotrimeric G proteins, but the mechanism underlying this quintessential molecular signaling event has remained unclear. Here we use atomic-level simulations to elucidate the nucleotide-release mechanism. We find that the G protein α subunit Ras and helical domains-previously observed to separate widely upon receptor binding to expose the nucleotide-binding site-separate spontaneously and frequently even in the absence of a receptor. Domain separation is necessary but not sufficient for rapid nucleotide release. Rather, receptors catalyze nucleotide release by favoring an internal structural rearrangement of the Ras domain that weakens its nucleotide affinity. We use double electron-electron resonance spectroscopy and protein engineering to confirm predictions of our computationally determined mechanism.
Copyright © 2015, American Association for the Advancement of Science.
Figures
References
-
- Oldham WM, Hamm HE. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat Rev Mol Cell Biol. 2008;9:60–71. - PubMed
-
- Sprang SR. G protein mechanisms: Insights from structural analysis. Ann Rev Biochem. 1997;66:639–678. - PubMed
-
- Venkatakrishnan AJ, Deupi X, Lebon G, Tate CG, Schertler GF, Babu MM. Molecular signatures of G-protein-coupled receptors. Nature. 2013;494:185–194. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
