

Deficiency Mutations of Alpha-1 Antitrypsin. Effects on Folding, Function, and Polymerization
- PMID: 26091018
- PMCID: PMC4742932
- DOI: 10.1165/rcmb.2015-0154OC
Deficiency Mutations of Alpha-1 Antitrypsin. Effects on Folding, Function, and Polymerization
Abstract
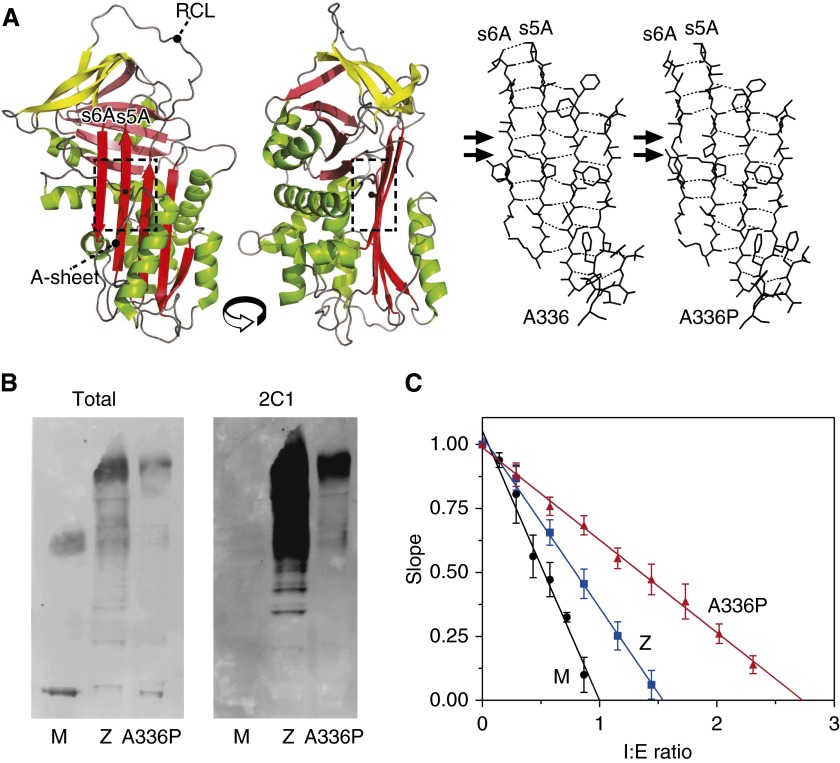
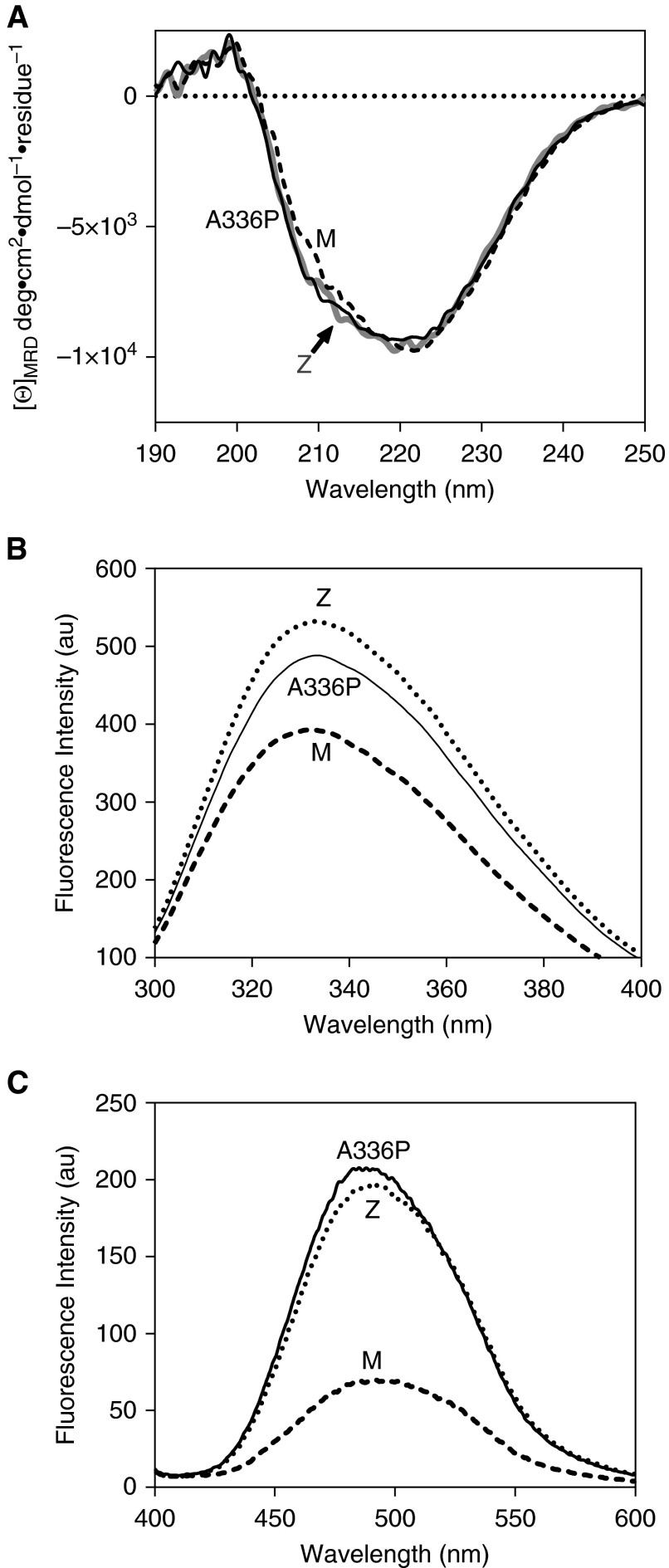
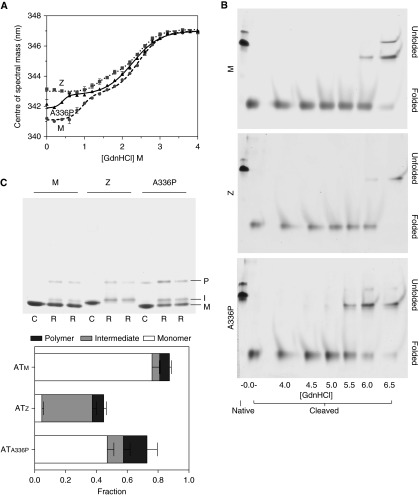
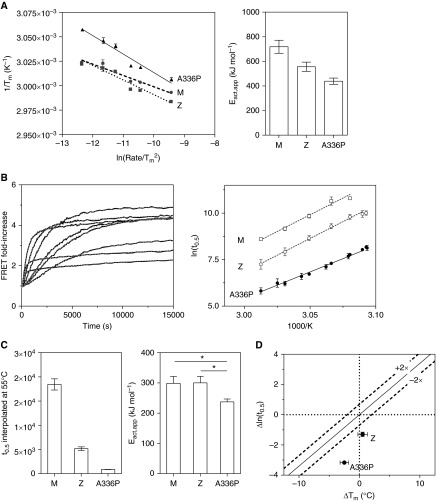
Misfolding, polymerization, and defective secretion of functional alpha-1 antitrypsin underlies the predisposition to severe liver and lung disease in alpha-1 antitrypsin deficiency. We have identified a novel (Ala336Pro, Baghdad) deficiency variant and characterized it relative to the wild-type (M) and Glu342Lys (Z) alleles. The index case is a homozygous individual of consanguineous parentage, with levels of circulating alpha-1 antitrypsin in the moderate deficiency range, but is a biochemical phenotype that could not be classified by standard methods. The majority of the protein was present as functionally inactive polymer, and the remaining monomer was 37% active relative to the wild-type protein. These factors combined indicate an 85 to 95% functional deficiency, similar to that seen with ZZ homozygotes. Biochemical, biophysical, and computational studies further defined the molecular basis of this deficiency. These studies demonstrated that native Ala336Pro alpha-1 antitrypsin could populate the polymerogenic intermediate-and therefore polymerize-more readily than either wild-type alpha-1 antitrypsin or the Z variant. In contrast, folding was far less impaired in Ala336Pro alpha-1 antitrypsin than in the Z variant. The data are consistent with a disparate contribution by the "breach" region and "shutter" region of strand 5A to folding and polymerization mechanisms. Moreover, the findings demonstrate that, in these variants, folding efficiency does not correlate directly with the tendency to polymerize in vitro or in vivo. They therefore differentiate generalized misfolding from polymerization tendencies in missense variants of alpha-1 antitrypsin. Clinically, they further support the need to quantify loss-of-function in alpha-1 antitrypsin deficiency to individualize patient care.
Keywords: alpha-1 antitrypsin deficiency; mutation; polymerization; serpinopathies; unfolding.
Figures
References
-
- Zaimidou S, van Baal S, Smith TD, Mitropoulos K, Ljujic M, Radojkovic D, Cotton RG, Patrinos GP. A1ATVar: a relational database of human SERPINA1 gene variants leading to α1-antitrypsin deficiency and application of the VariVis software. Hum Mutat. 2009;30:308–313. - PubMed
-
- Brantly ML, Wittes JT, Vogelmeier CF, Hubbard RC, Fells GA, Crystal RG. Use of a highly purified alpha 1-antitrypsin standard to establish ranges for the common normal and deficient alpha 1-antitrypsin phenotypes. Chest. 1991;100:703–708. - PubMed
-
- Allan PC, Harley RA, Talamo RC. A new method for determination of alpha-1-antitrypsin phenotype using isoelectric focusing on polyacrylamide gel slabs. Am J Clin Pathol. 1974;62:732–739. - PubMed
-
- Sveger T. Liver disease in alpha1-antitrypsin deficiency detected by screening of 200,000 infants. N Engl J Med. 1976;294:1316–1321. - PubMed
-
- Silverman EK, Miletich JP, Pierce JA, Sherman LA, Endicott SK, Broze GJ, Jr, Campbell EJ. Alpha-1-antitrypsin deficiency: high prevalence in the St. Louis area determined by direct population screening. Am Rev Respir Dis. 1989;140:961–966. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
