

Embryonic Origin of Postnatal Neural Stem Cells
- PMID: 26091041
- PMCID: PMC4475276
- DOI: 10.1016/j.cell.2015.05.041
Embryonic Origin of Postnatal Neural Stem Cells
Abstract
Adult neural stem/progenitor (B1) cells within the walls of the lateral ventricles generate different types of neurons for the olfactory bulb (OB). The location of B1 cells determines the types of OB neurons they generate. Here we show that the majority of mouse B1 cell precursors are produced between embryonic days (E) 13.5 and 15.5 and remain largely quiescent until they become reactivated postnatally. Using a retroviral library carrying over 100,000 genetic tags, we found that B1 cells share a common progenitor with embryonic cells of the cortex, striatum, and septum, but this lineage relationship is lost before E15.5. The regional specification of B1 cells is evident as early as E11.5 and is spatially linked to the production of neurons that populate different areas of the forebrain. This study reveals an early embryonic regional specification of postnatal neural stem cells and the lineage relationship between them and embryonic progenitor cells.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

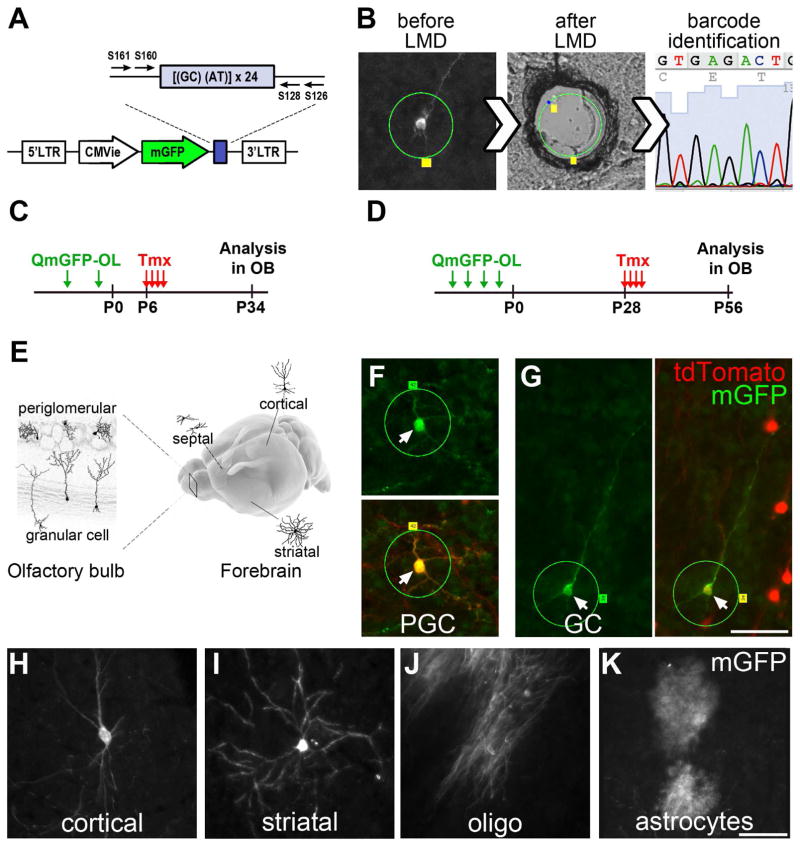
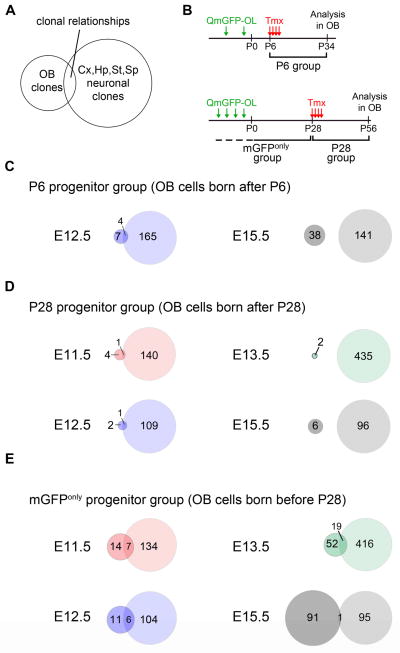
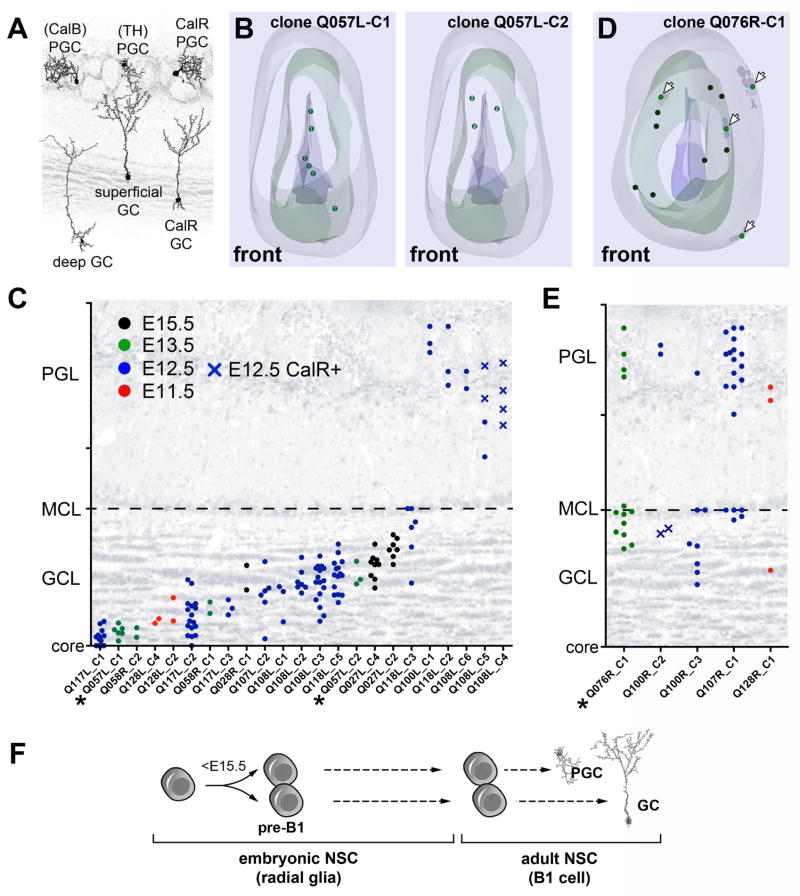
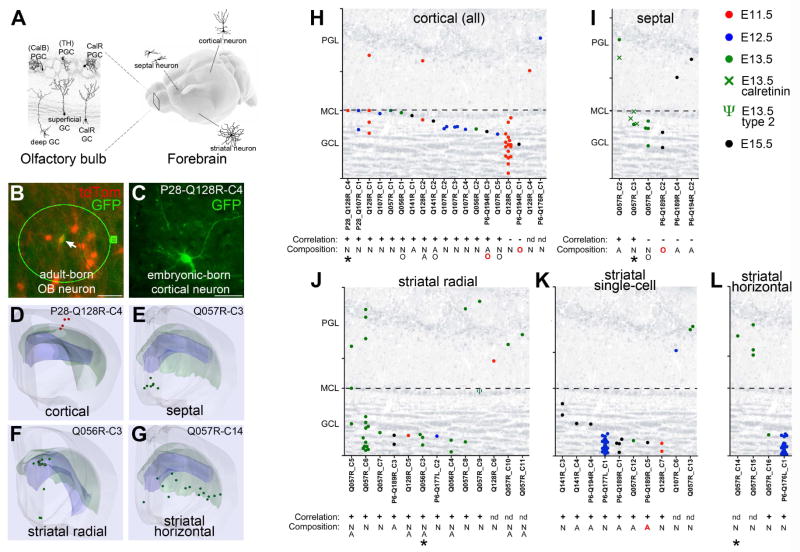
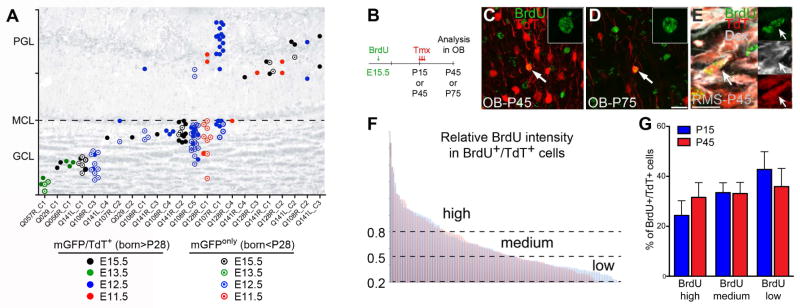
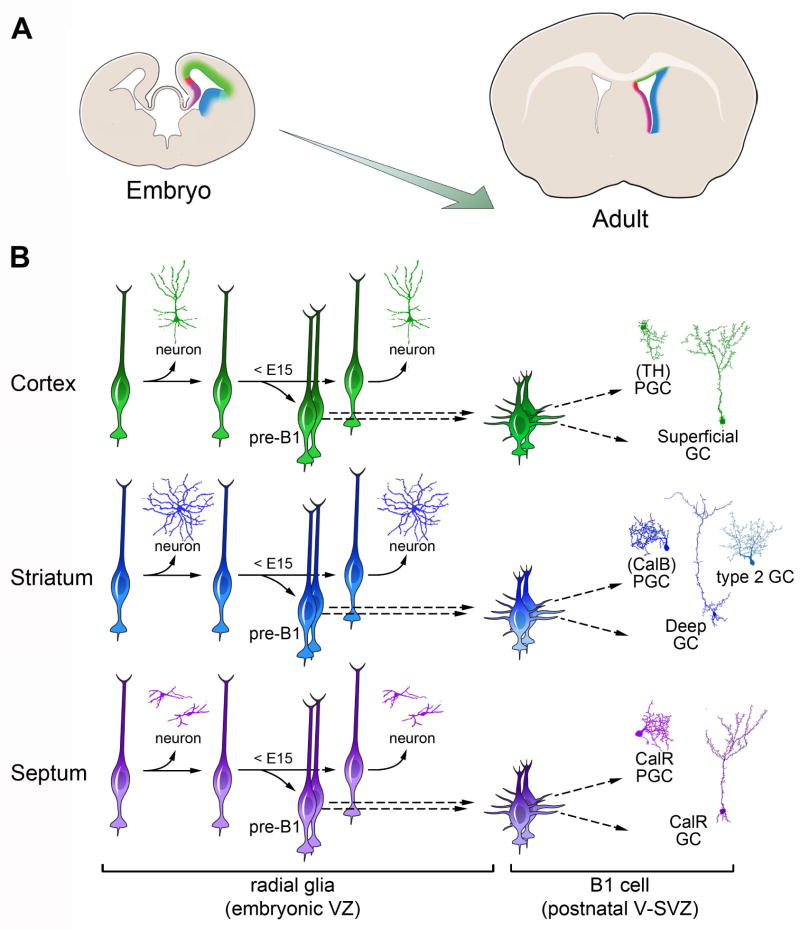
Comment in
-
Finding the Roots of Adult Neurogenesis.Cell. 2015 Jun 18;161(7):1500-2. doi: 10.1016/j.cell.2015.06.010. Cell. 2015. PMID: 26091033 Free PMC article.
References
-
- Alvarez-Buylla A, García-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. - PubMed
-
- Alvarez-Buylla A, Kohwi M, Nguyen TM, Merkle FT. The heterogeneity of adult neural stem cells and the emerging complexity of their niche. Cold Spring Harb Symp Quant Biol. 2008;73:357–365. - PubMed
-
- Bayer SA. 3H-thymidine-radiographic studies of neurogenesis in the rat olfactory bulb. Exp Brain Res. 1983;50:329–340. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
