

Cancer-associated ASXL1 mutations may act as gain-of-function mutations of the ASXL1-BAP1 complex
- PMID: 26095772
- PMCID: PMC4557297
- DOI: 10.1038/ncomms8307
Cancer-associated ASXL1 mutations may act as gain-of-function mutations of the ASXL1-BAP1 complex
Abstract
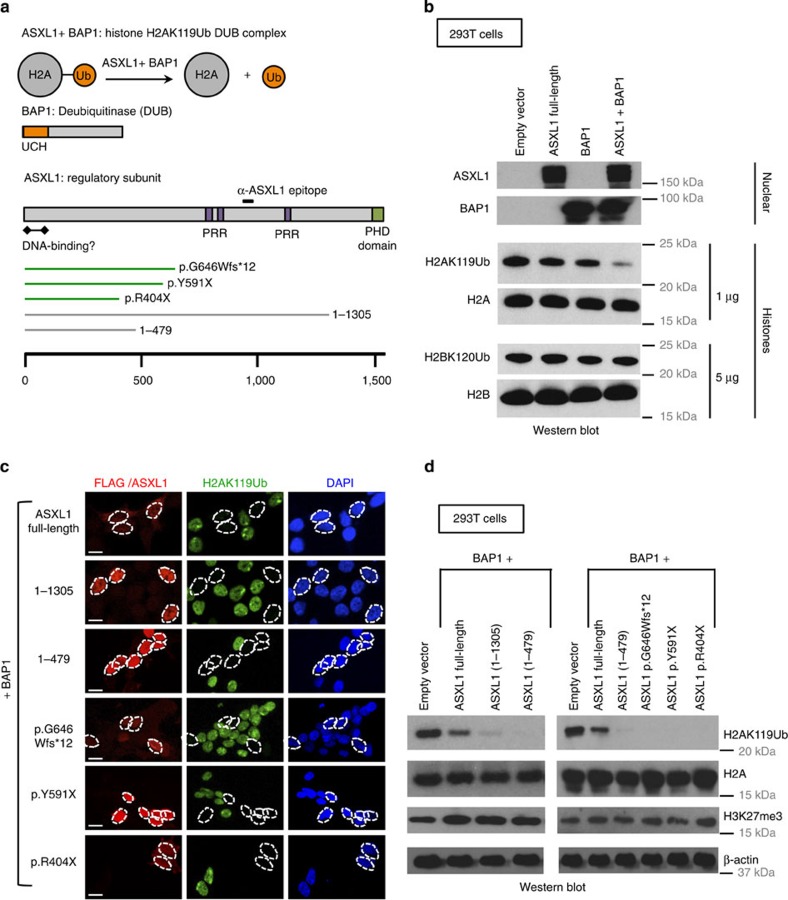
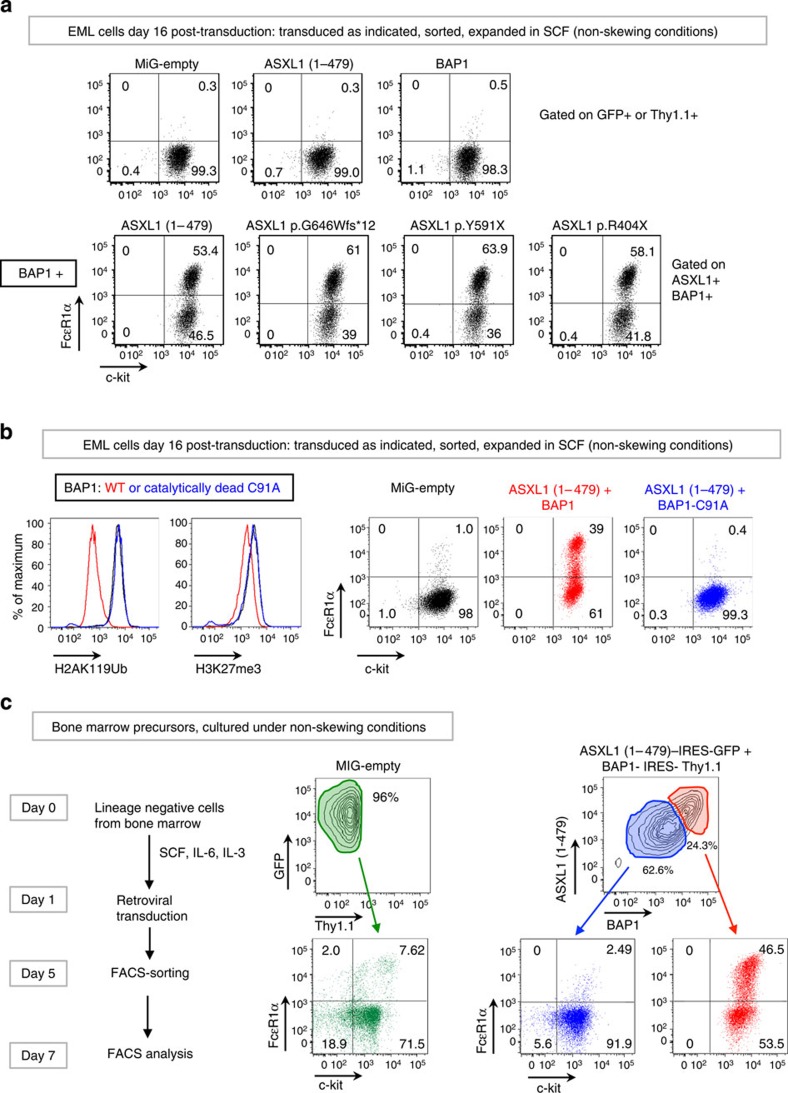
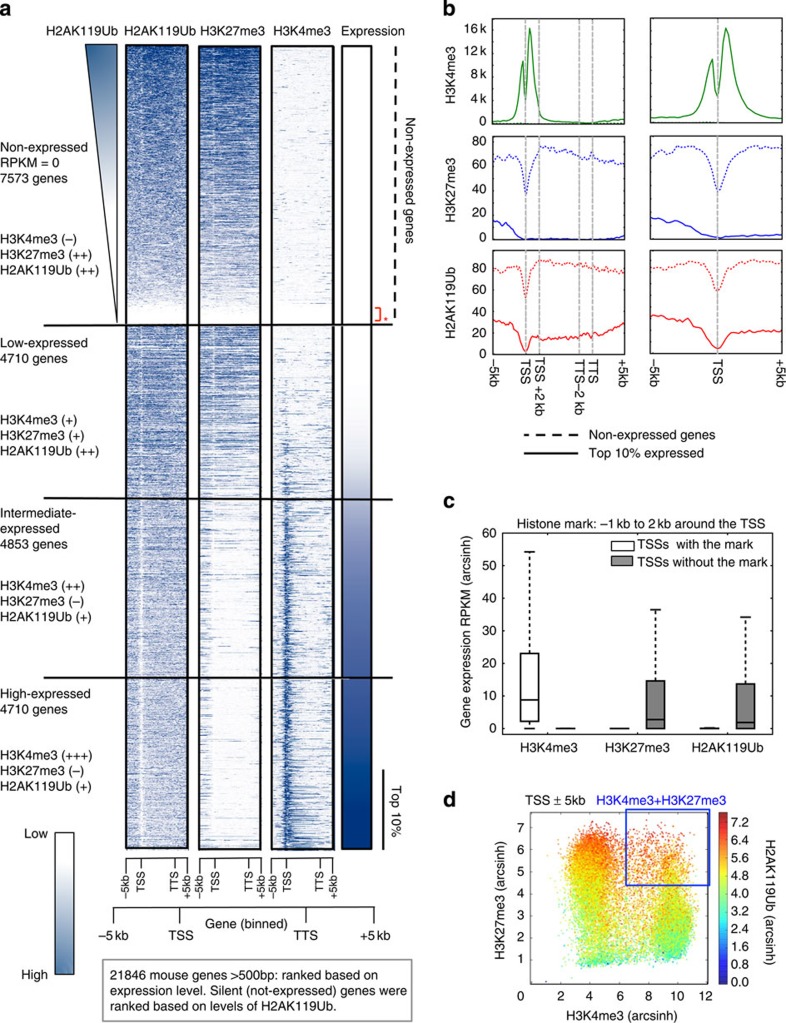
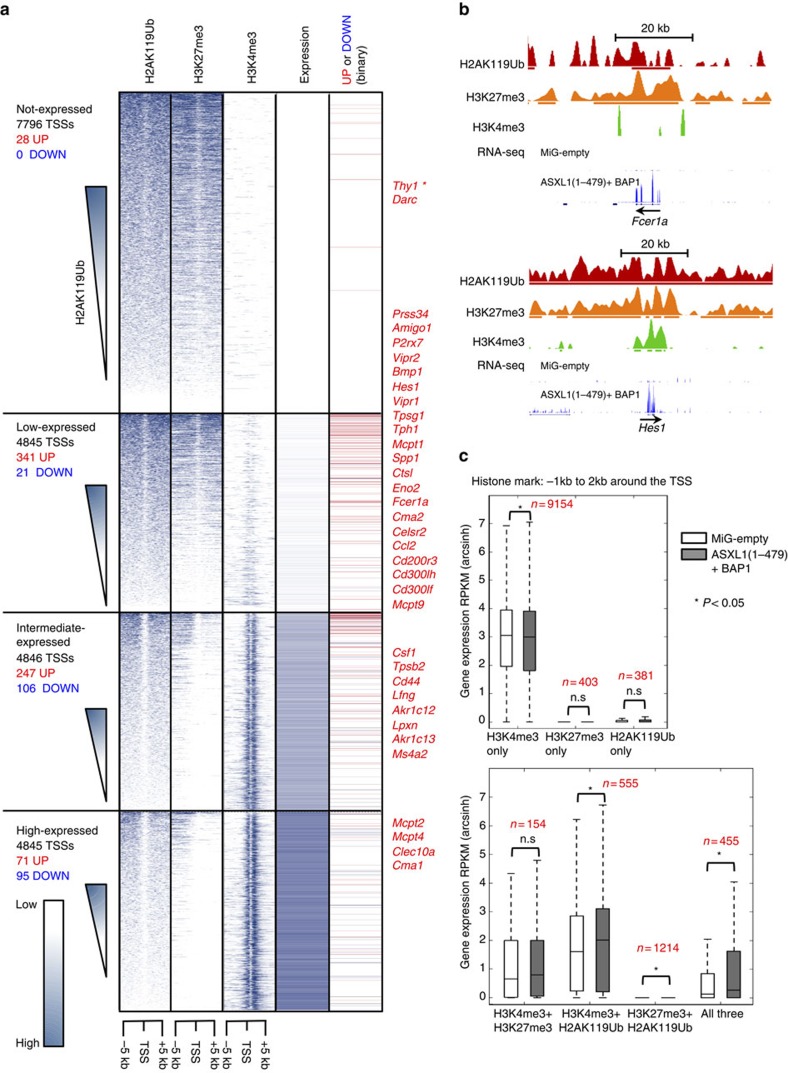
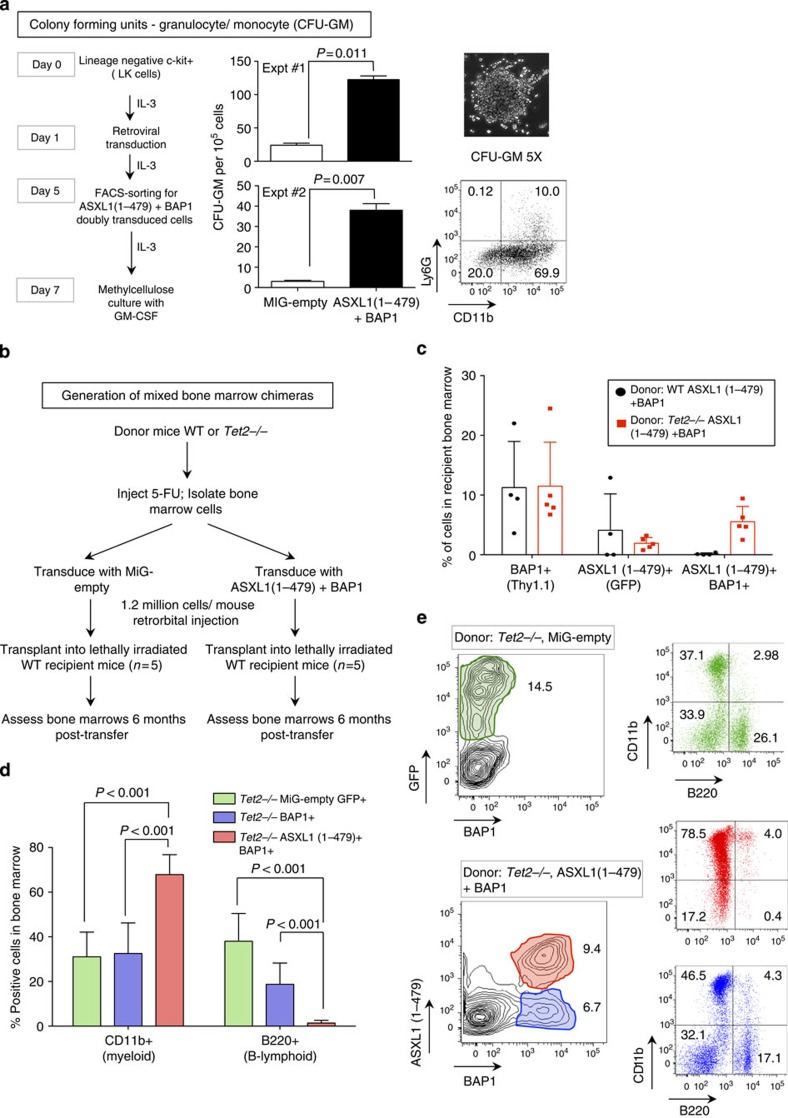
ASXL1 is the obligate regulatory subunit of a deubiquitinase complex whose catalytic subunit is BAP1. Heterozygous mutations of ASXL1 that result in premature truncations are frequent in myeloid leukemias and Bohring-Opitz syndrome. Here we demonstrate that ASXL1 truncations confer enhanced activity on the ASXL1-BAP1 complex. Stable expression of truncated, hyperactive ASXL1-BAP1 complexes in a haematopoietic precursor cell line results in global erasure of H2AK119Ub, striking depletion of H3K27me3, selective upregulation of a subset of genes whose promoters are marked by both H2AK119Ub and H3K4me3, and spontaneous differentiation to the mast cell lineage. These outcomes require the catalytic activity of BAP1, indicating that they are downstream consequences of H2AK119Ub erasure. In bone marrow precursors, expression of truncated ASXL1-BAP1 complex cooperates with TET2 loss-of-function to increase differentiation to the myeloid lineage in vivo. Our data raise the possibility that ASXL1 truncation mutations confer gain-of-function on the ASXL-BAP1 complex.
Figures

References
-
- Jurgens G. A group of genes controlling expression of the bithorax complex. Nature 316, 1985 (1985).
-
- Wang H. et al. Role of histone H2A ubiquitination in Polycomb silencing. Nature 431, 873–878 (2004). - PubMed
-
- Muller J. et al. Histone methyltransferase activity of a Drosophila Polycomb group repressor complex. Cell 111, 197–208 (2002). - PubMed
-
- Czermin B. et al. Drosophila enhancer of Zeste/ESC complexes have a histone H3 methyltransferase activity that marks chromosomal Polycomb sites. Cell 111, 185–196 (2002). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
