

Comprehensive translational control of tyrosine kinase expression by upstream open reading frames
- PMID: 26096937
- PMCID: PMC4820681
- DOI: 10.1038/onc.2015.233
Comprehensive translational control of tyrosine kinase expression by upstream open reading frames
Abstract
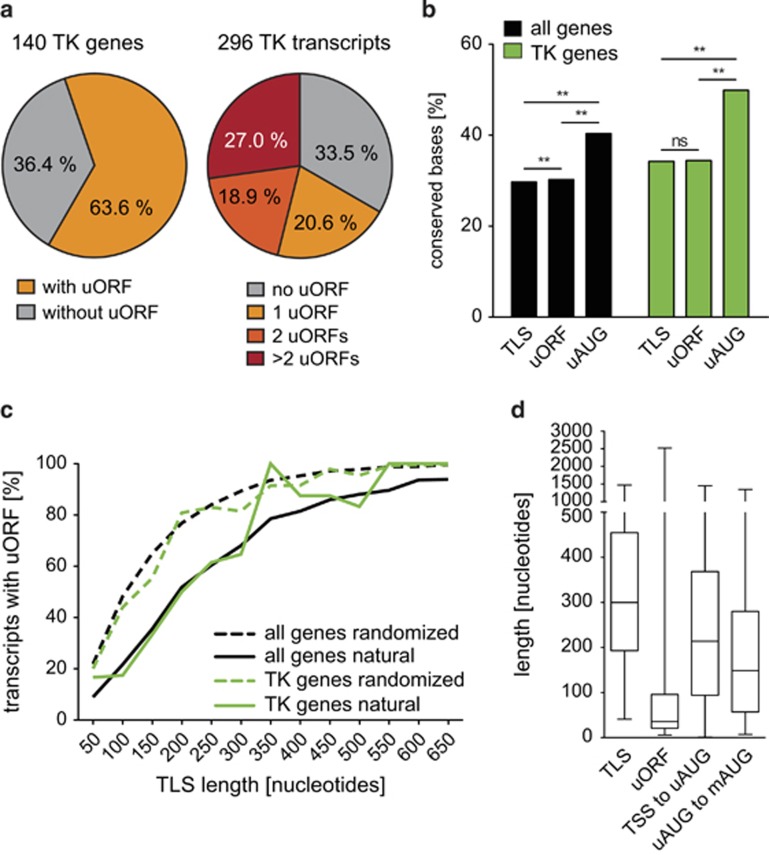
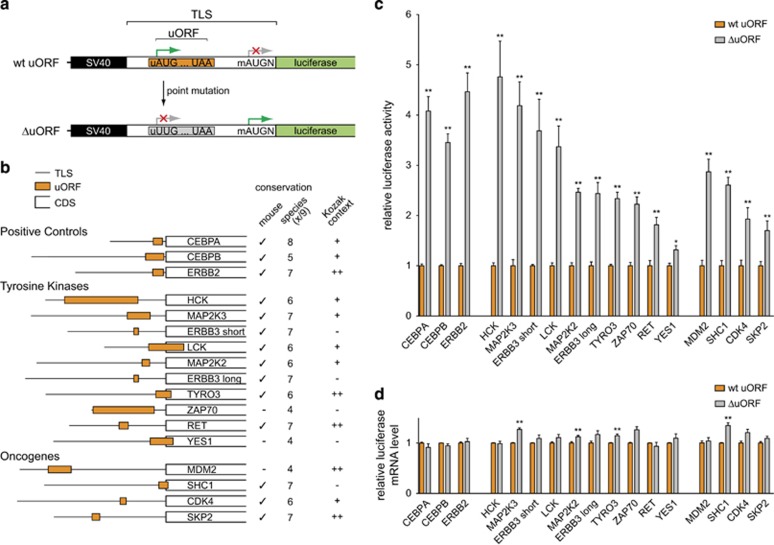
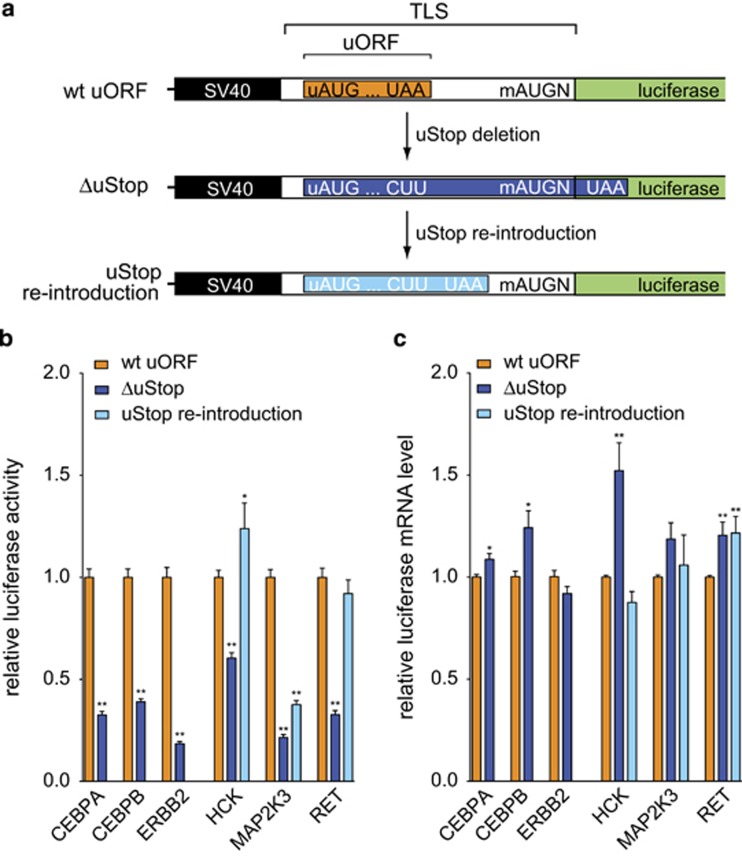
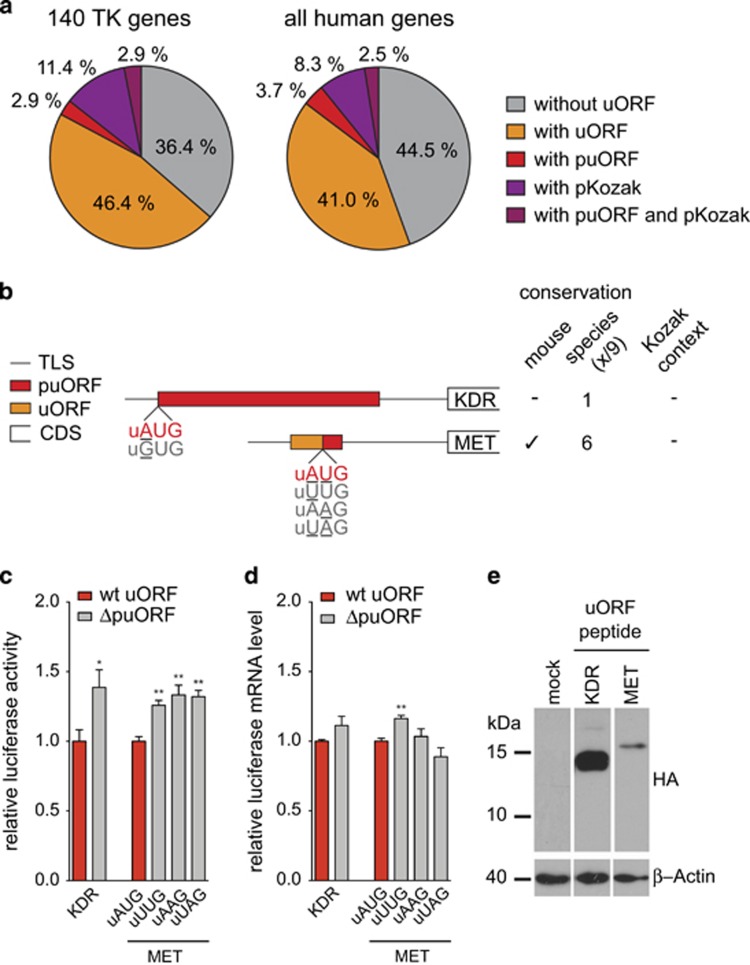
Post-transcriptional control has emerged as a major regulatory event in gene expression and often occurs at the level of translation initiation. Although overexpression or constitutive activation of tyrosine kinases (TKs) through gene amplification, translocation or mutation are well-characterized oncogenic events, current knowledge about translational mechanisms of TK activation is scarce. Here, we report the presence of translational cis-regulatory upstream open reading frames (uORFs) in the majority of transcript leader sequences of human TK mRNAs. Genetic ablation of uORF initiation codons in TK transcripts resulted in enhanced translation of the associated downstream main protein-coding sequences (CDSs) in all cases studied. Similarly, experimental removal of uORF start codons in additional non-TK proto-oncogenes, and naturally occurring loss-of-uORF alleles of the c-met proto-oncogene (MET) and the kinase insert domain receptor (KDR), was associated with increased CDS translation. Based on genome-wide sequence analyses we identified polymorphisms in 15.9% of all human genes affecting uORF initiation codons, associated Kozak consensus sequences or uORF-related termination codons. Together, these data suggest a comprehensive role of uORF-mediated translational control and delineate how aberrant induction of proto-oncogenes through loss-of-function mutations at uORF initiation codons may be involved in the etiology of cancer. We provide a detailed map of uORFs across the human genome to stimulate future research on the pathogenic role of uORFs.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
