

Subtype-specific plasticity of inhibitory circuits in motor cortex during motor learning
- PMID: 26098758
- PMCID: PMC4519436
- DOI: 10.1038/nn.4049
Subtype-specific plasticity of inhibitory circuits in motor cortex during motor learning
Abstract
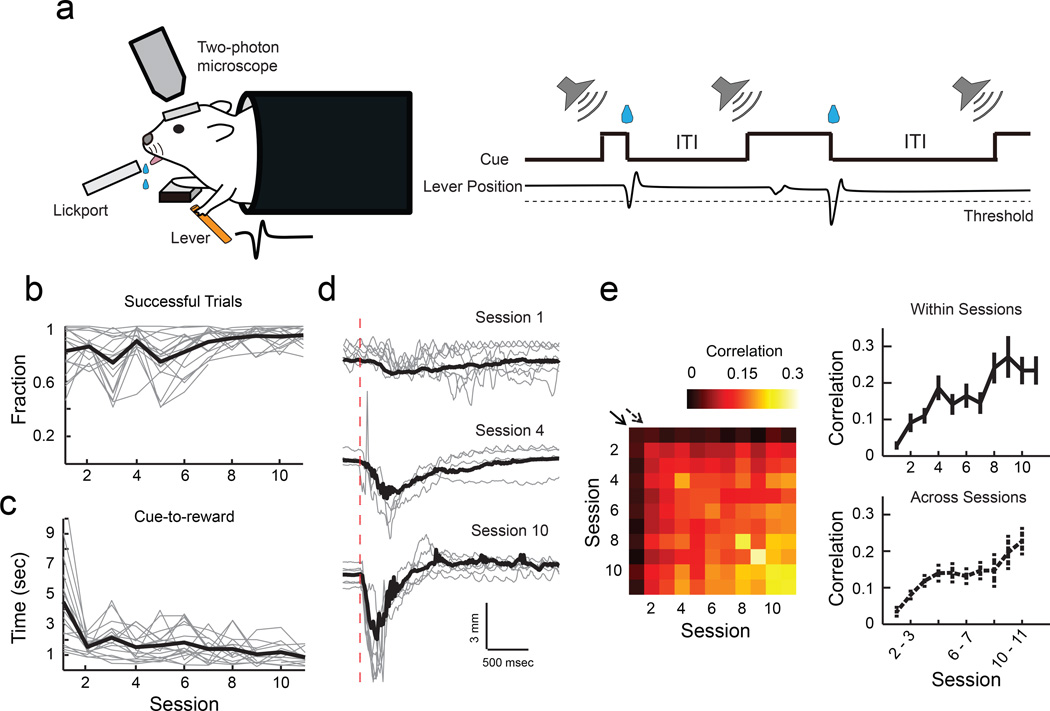
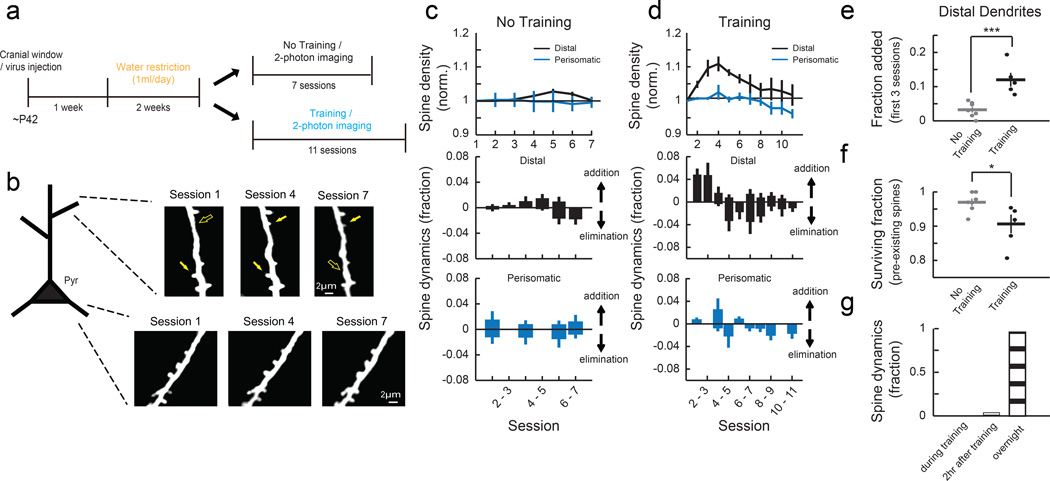
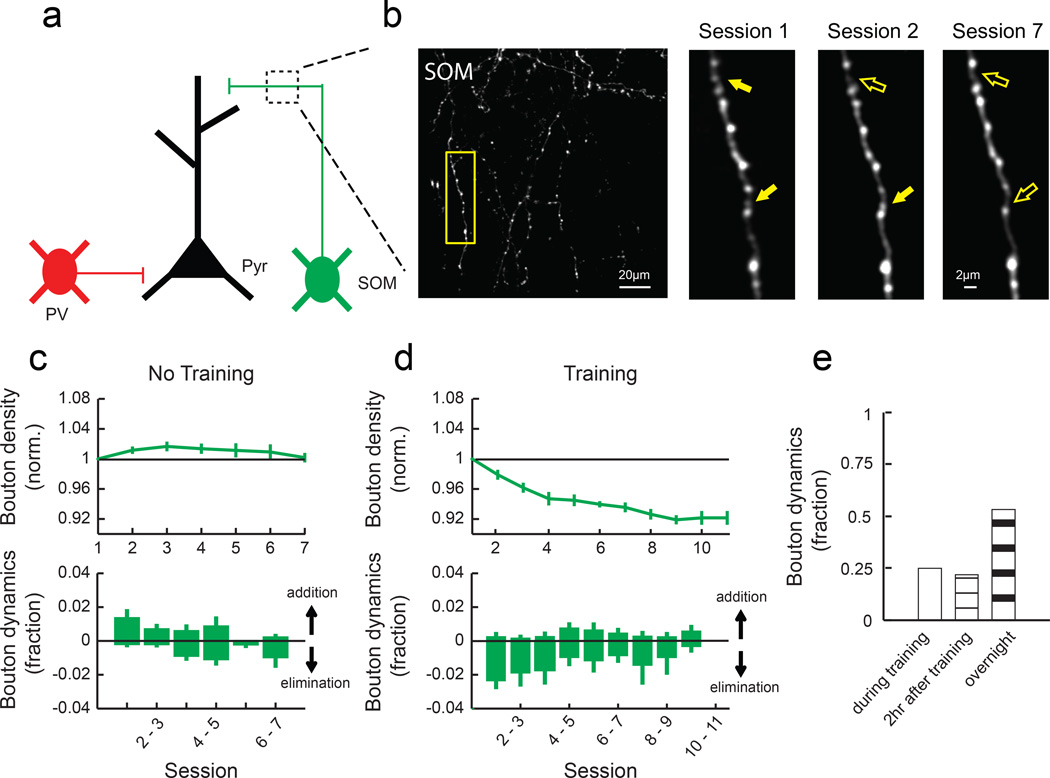
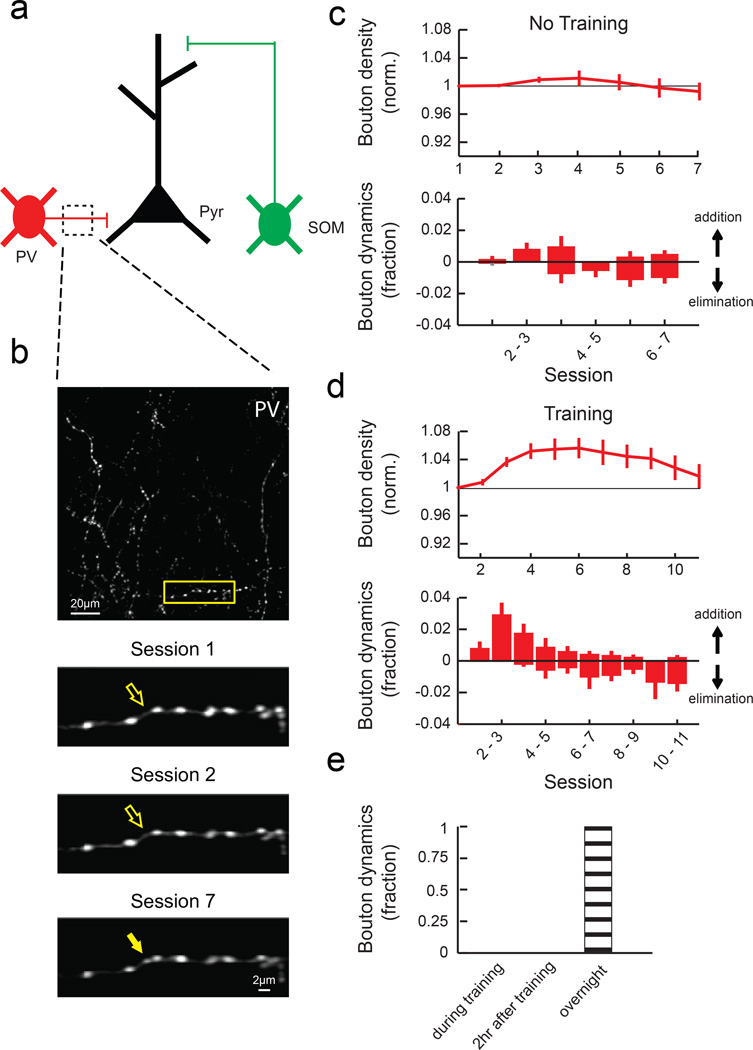
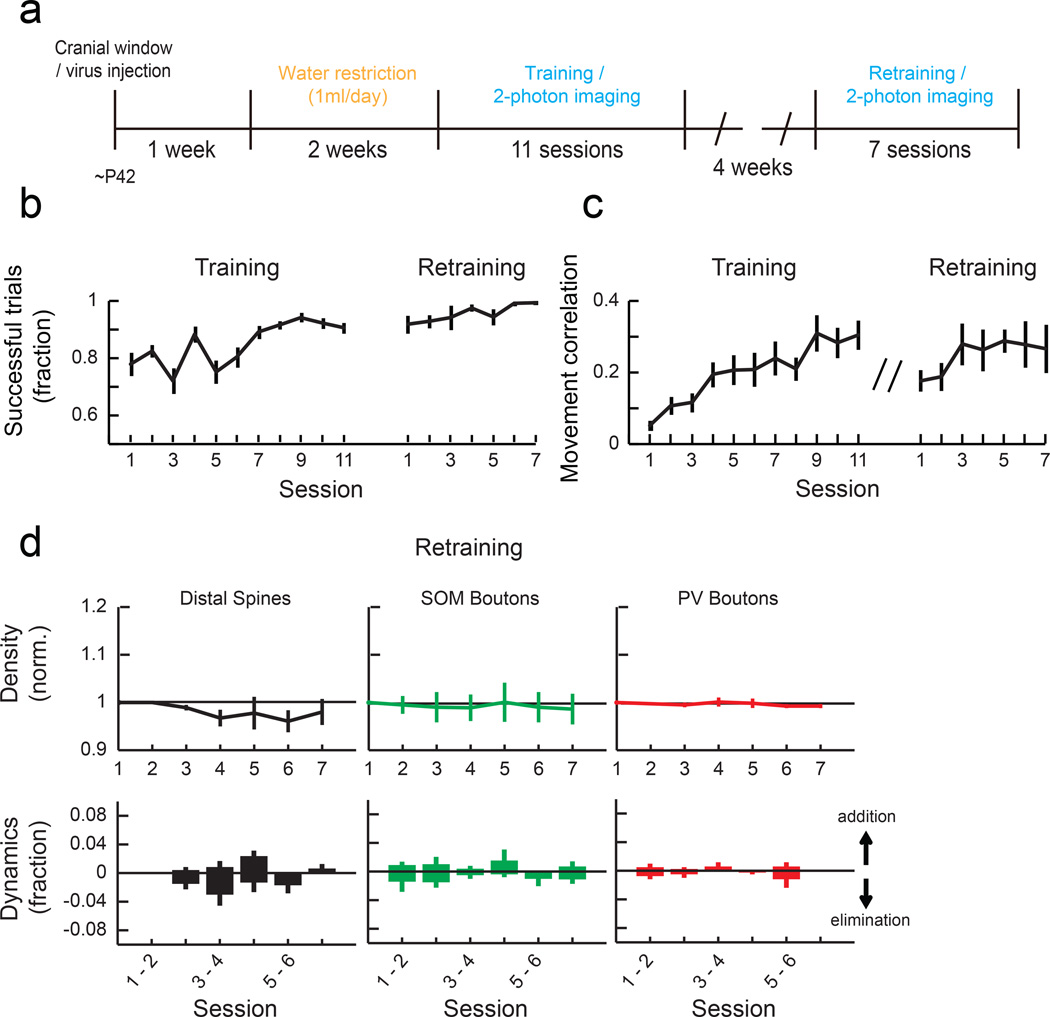
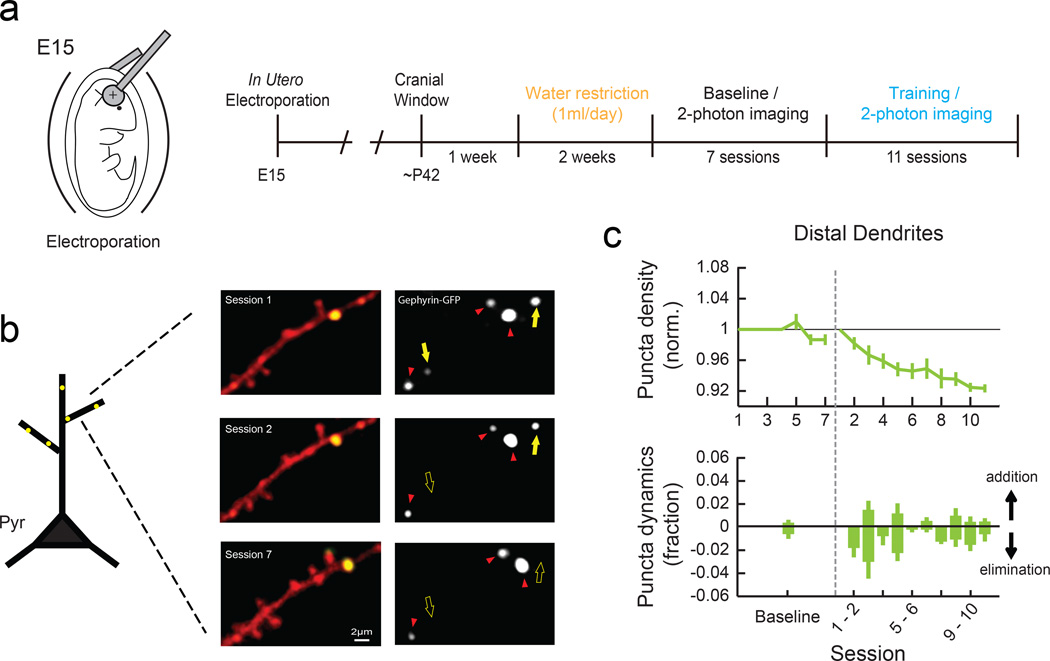
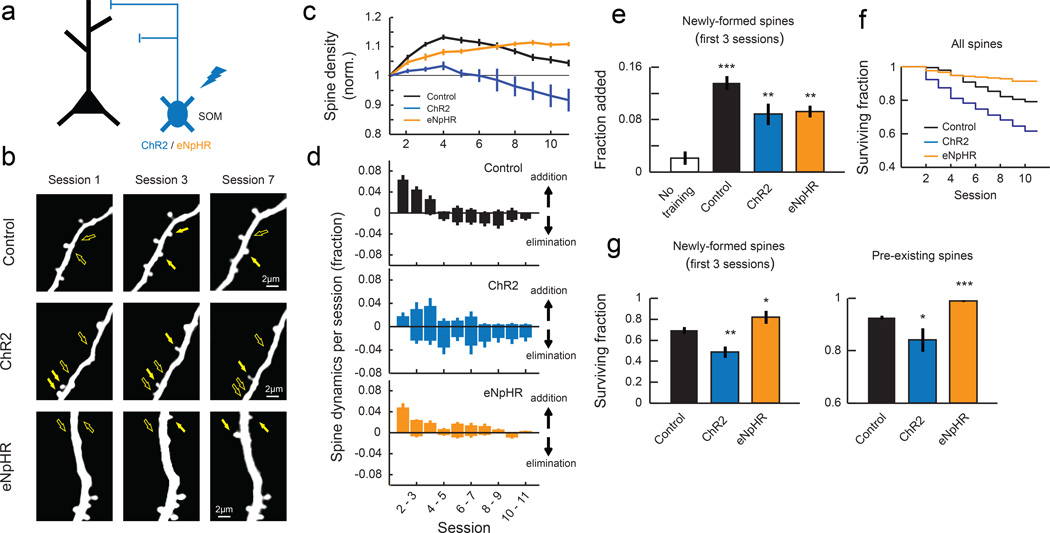
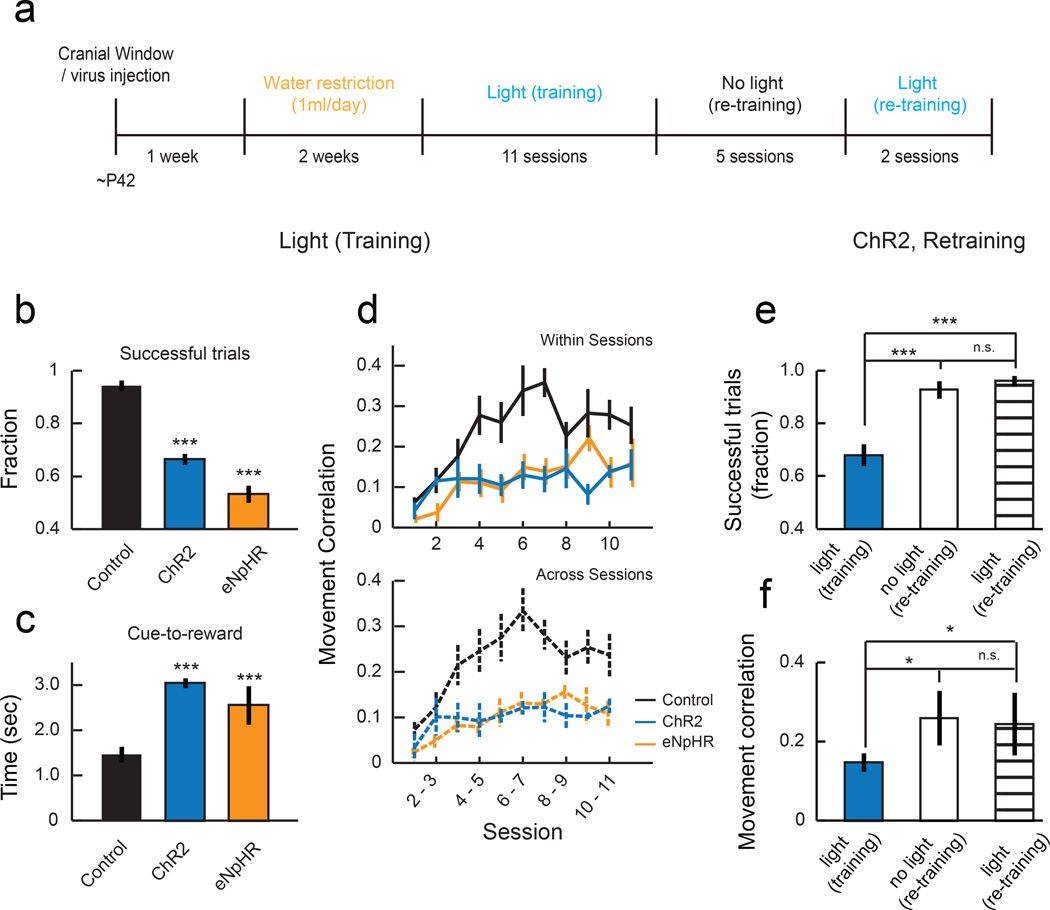
Motor skill learning induces long-lasting reorganization of dendritic spines, principal sites of excitatory synapses, in the motor cortex. However, mechanisms that regulate these excitatory synaptic changes remain poorly understood. Here, using in vivo two-photon imaging in awake mice, we found that learning-induced spine reorganization of layer (L) 2/3 excitatory neurons occurs in the distal branches of their apical dendrites in L1 but not in the perisomatic dendrites. This compartment-specific spine reorganization coincided with subtype-specific plasticity of local inhibitory circuits. Somatostatin-expressing inhibitory neurons (SOM-INs), which mainly inhibit distal dendrites of excitatory neurons, showed a decrease in axonal boutons immediately after the training began, whereas parvalbumin-expressing inhibitory neurons (PV-INs), which mainly inhibit perisomatic regions of excitatory neurons, exhibited a gradual increase in axonal boutons during training. Optogenetic enhancement and suppression of SOM-IN activity during training destabilized and hyperstabilized spines, respectively, and both manipulations impaired the learning of stereotyped movements. Our results identify SOM inhibition of distal dendrites as a key regulator of learning-related changes in excitatory synapses and the acquisition of motor skills.
Figures
Comment in
-
Removing synaptic brakes on learning.Nat Neurosci. 2015 Aug;18(8):1062-4. doi: 10.1038/nn.4073. Nat Neurosci. 2015. PMID: 26216460 No abstract available.
References
-
- Peters AJ, Chen SX, Komiyama T. Emergence of reproducible spatiotemporal activity during motor learning. Nature. 2014;510:263–267. - PubMed
-
- Costa RM, Cohen D, Nicolelis MA. Differential corticostriatal plasticity during fast and slow motor skill learning in mice. Curr Biol. 2004;14:1124–1134. - PubMed
-
- Rioult-Pedotti MS, Friedman D, Donoghue JP. Learning-induced LTP in neocortex. Science. 2000;290:533–536. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
