

Shell neurons of the master circadian clock coordinate the phase of tissue clocks throughout the brain and body
- PMID: 26099272
- PMCID: PMC4489020
- DOI: 10.1186/s12915-015-0157-x
Shell neurons of the master circadian clock coordinate the phase of tissue clocks throughout the brain and body
Abstract
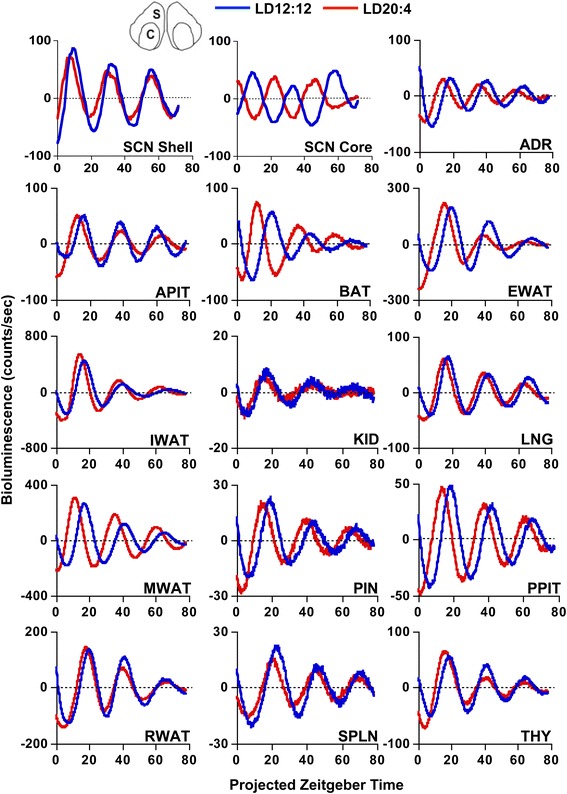
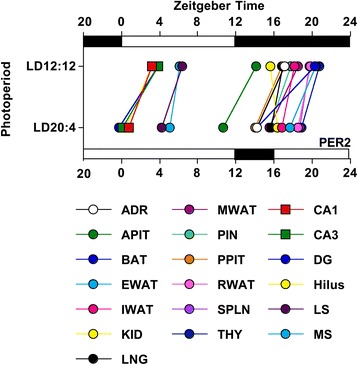
Background: Daily rhythms in mammals are programmed by a master clock in the suprachiasmatic nucleus (SCN). The SCN contains two main compartments (shell and core), but the role of each region in system-level coordination remains ill defined. Herein, we use a functional assay to investigate how downstream tissues interpret region-specific outputs by using in vivo exposure to long day photoperiods to temporally dissociate the SCN. We then analyze resulting changes in the rhythms of clocks located throughout the brain and body to examine whether they maintain phase synchrony with the SCN shell or core.
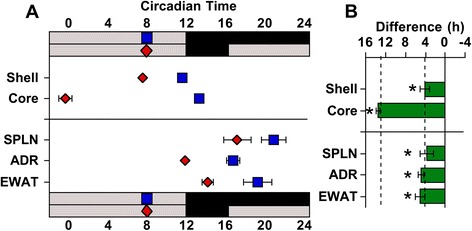
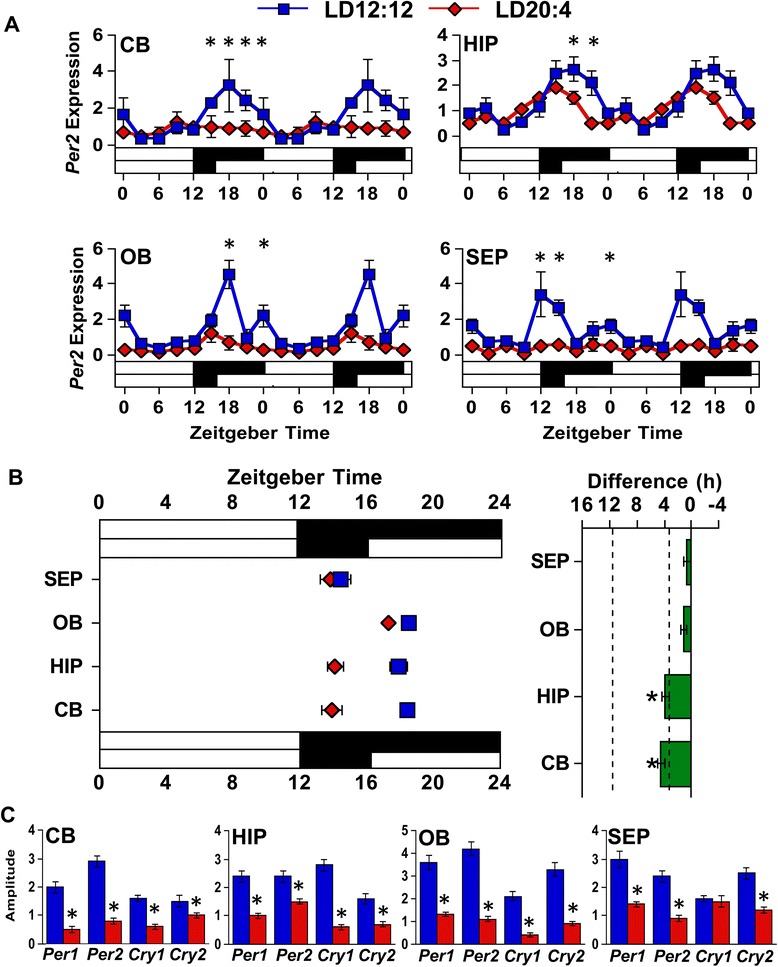
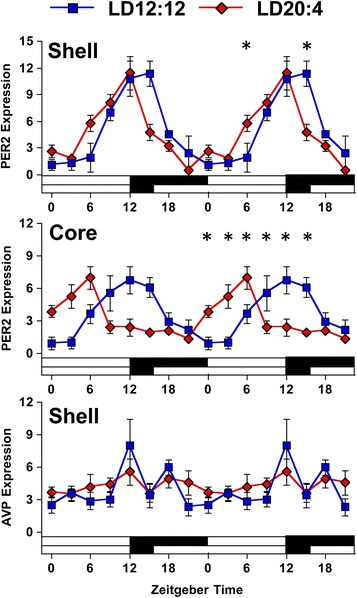
Results: Nearly all of the 17 tissues examined in the brain and body maintain phase synchrony with the SCN shell, but not the SCN core, which indicates that downstream oscillators are set by cues controlled specifically by the SCN shell. Interestingly, we also found that SCN dissociation diminished the amplitude of rhythms in core clock gene and protein expression in brain tissues by 50-75 %, which suggests that light-driven changes in the functional organization of the SCN markedly influence the strength of rhythms in downstream tissues.
Conclusions: Overall, our results reveal that body clocks receive time-of-day cues specifically from the SCN shell, which may be an adaptive design principle that serves to maintain system-level phase relationships in a changing environment. Further, we demonstrate that lighting conditions alter the amplitude of the molecular clock in downstream tissues, which uncovers a new form of plasticity that may contribute to seasonal changes in physiology and behavior.
Figures


References
Publication types
MeSH terms
Grants and funding
- R01 EY022216/EY/NEI NIH HHS/United States
- U54 NS083932/NS/NINDS NIH HHS/United States
- SC1 GM112567/GM/NIGMS NIH HHS/United States
- 8G12MD007602/MD/NIMHD NIH HHS/United States
- U54NS060659/NS/NINDS NIH HHS/United States
- R37 DK035254/DK/NIDDK NIH HHS/United States
- R01 NS078410/NS/NINDS NIH HHS/United States
- S21MD000101/MD/NIMHD NIH HHS/United States
- R01 DK078358/DK/NIDDK NIH HHS/United States
- R01EY022216/EY/NEI NIH HHS/United States
- U54NS083932/NS/NINDS NIH HHS/United States
- S21 MD000101/MD/NIMHD NIH HHS/United States
- U54 NS060659/NS/NINDS NIH HHS/United States
- F32 NS071935/NS/NINDS NIH HHS/United States
- R01NS078410/NS/NINDS NIH HHS/United States
- F32NS071935/NS/NINDS NIH HHS/United States
- G12 MD007602/MD/NIMHD NIH HHS/United States
- R37DK35254/DK/NIDDK NIH HHS/United States
- R01 DK035254/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
