

The bacterial tubulin FtsZ requires its intrinsically disordered linker to direct robust cell wall construction
- PMID: 26099469
- PMCID: PMC4532373
- DOI: 10.1038/ncomms8281
The bacterial tubulin FtsZ requires its intrinsically disordered linker to direct robust cell wall construction
Abstract
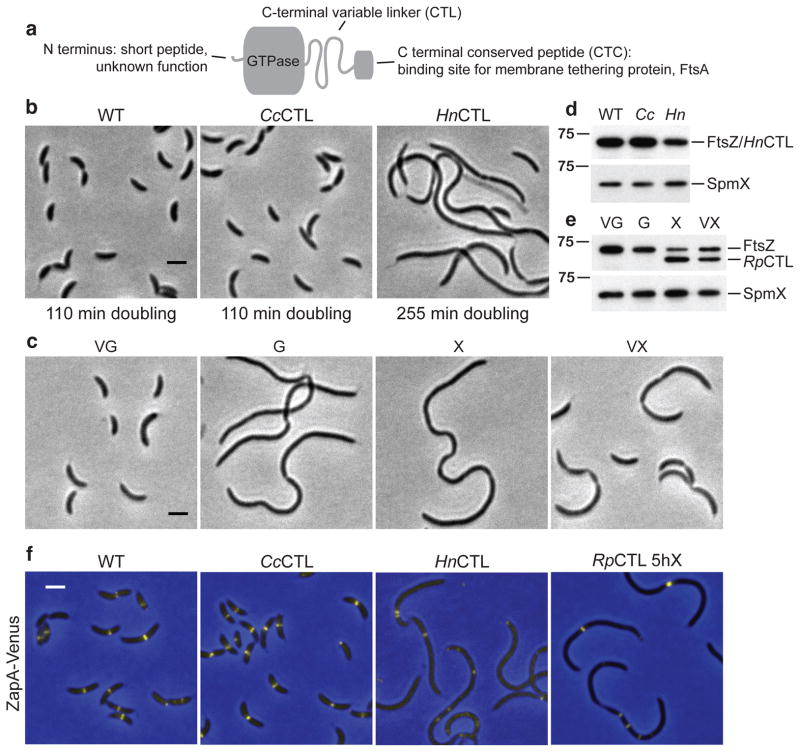
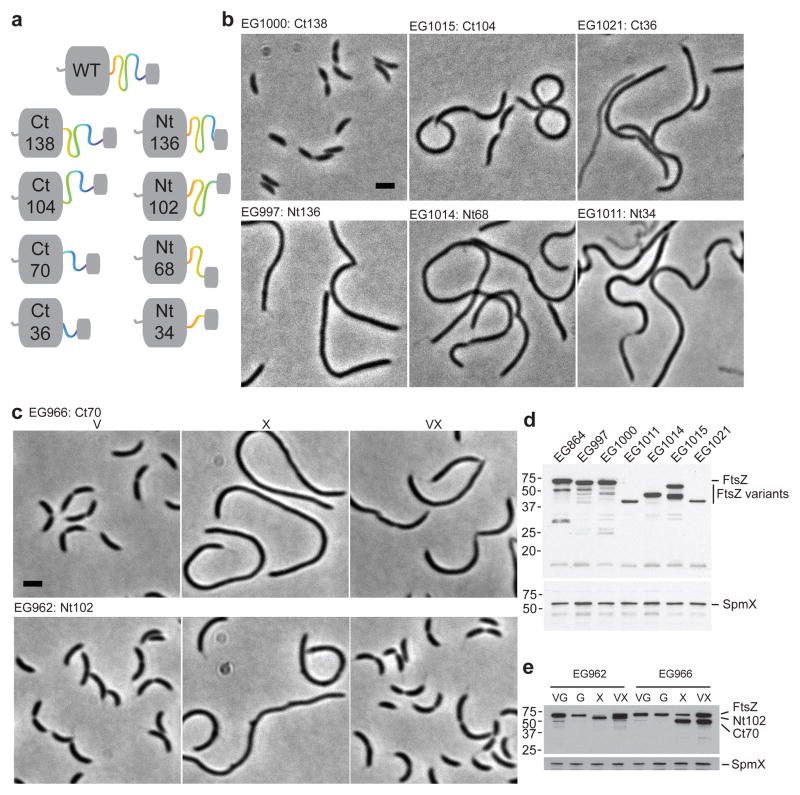
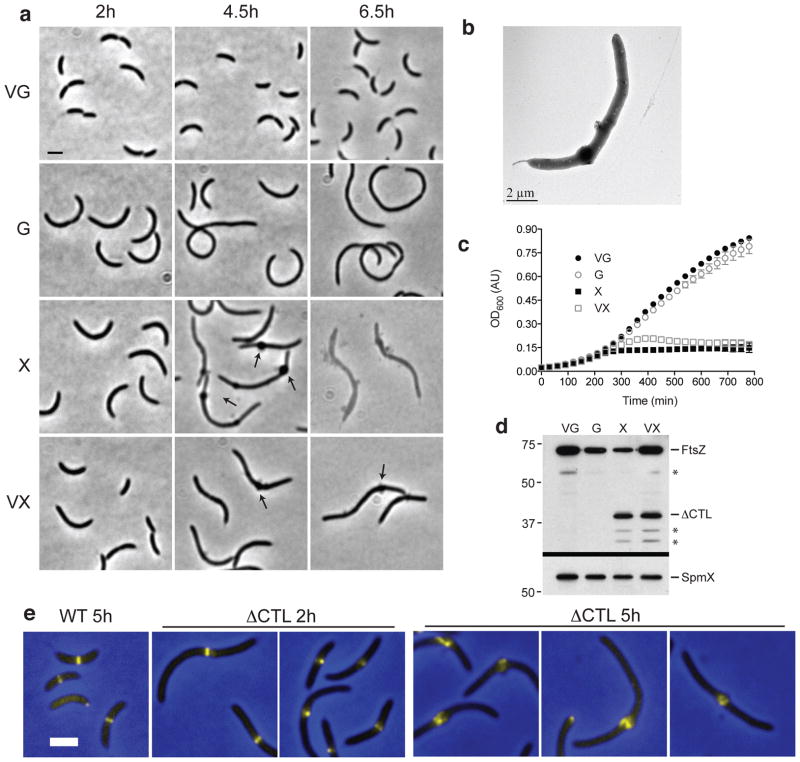
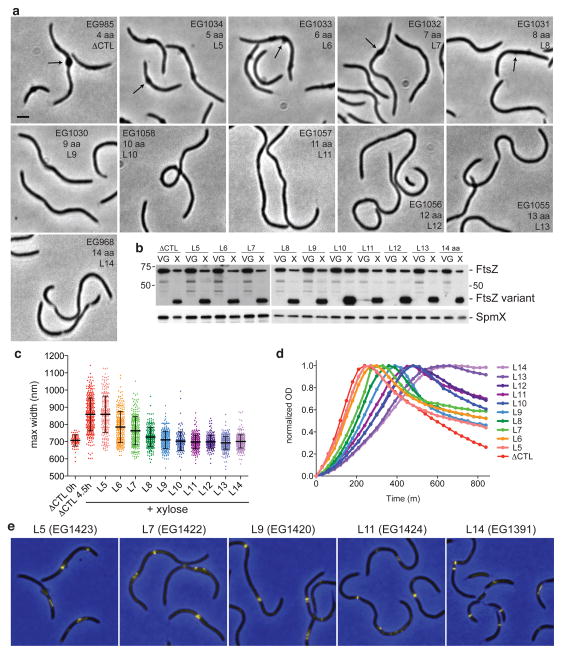
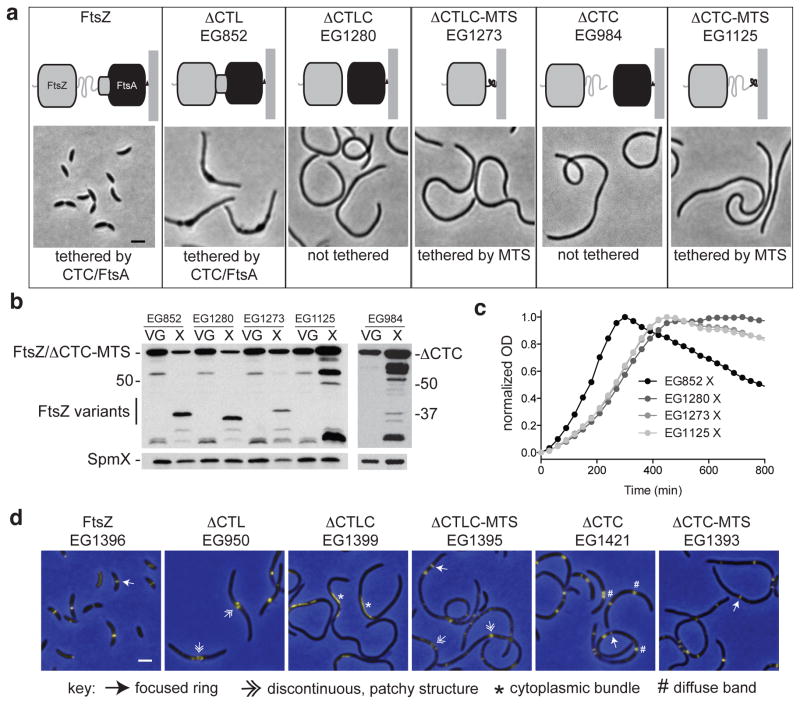
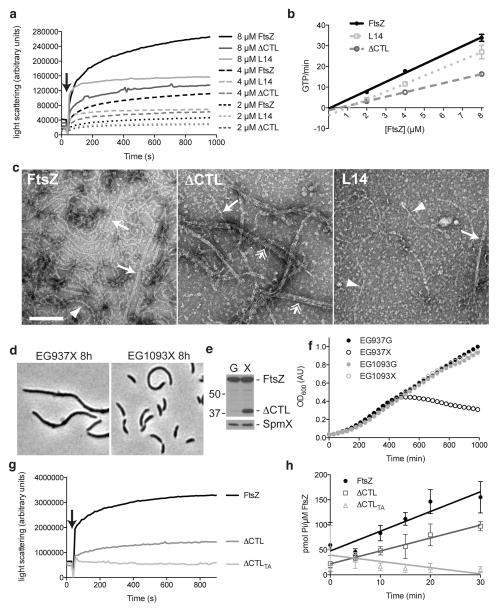
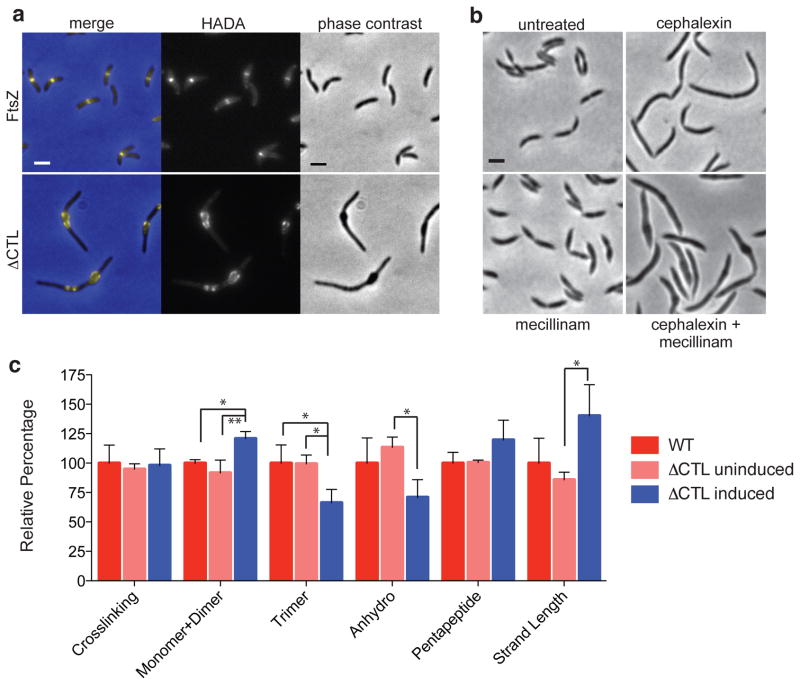
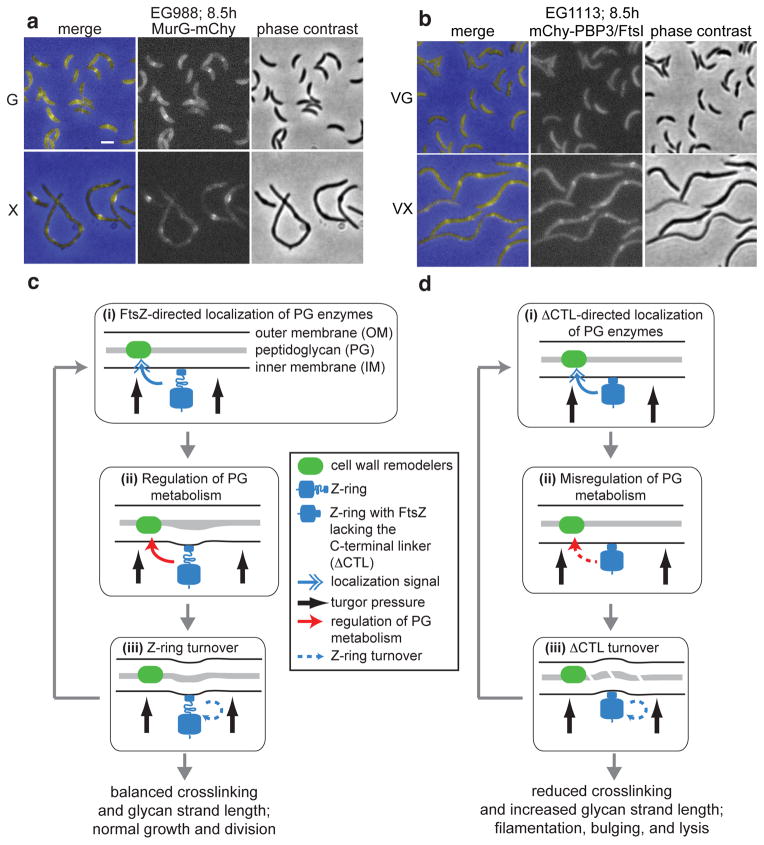
The bacterial GTPase FtsZ forms a cytokinetic ring at midcell, recruits the division machinery and orchestrates membrane and peptidoglycan cell wall invagination. However, the mechanism for FtsZ regulation of peptidoglycan metabolism is unknown. The FtsZ GTPase domain is separated from its membrane-anchoring C-terminal conserved (CTC) peptide by a disordered C-terminal linker (CTL). Here we investigate CTL function in Caulobacter crescentus. Strikingly, production of FtsZ lacking the CTL (ΔCTL) is lethal: cells become filamentous, form envelope bulges and lyse, resembling treatment with β-lactam antibiotics. This phenotype is produced by FtsZ polymers bearing the CTC and a CTL shorter than 14 residues. Peptidoglycan synthesis still occurs downstream of ΔCTL; however, cells expressing ΔCTL exhibit reduced peptidoglycan crosslinking and longer glycan strands than wild type. Importantly, midcell proteins are still recruited to sites of ΔCTL assembly. We propose that FtsZ regulates peptidoglycan metabolism through a CTL-dependent mechanism that extends beyond simple protein recruitment.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Wang S, Shaevitz JW. The mechanics of shape in prokaryotes. Front Biosci (Schol Ed) 2013;5:564–574. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
