

Toxicity and SidJ-Mediated Suppression of Toxicity Require Distinct Regions in the SidE Family of Legionella pneumophila Effectors
- PMID: 26099583
- PMCID: PMC4534637
- DOI: 10.1128/IAI.00497-15
Toxicity and SidJ-Mediated Suppression of Toxicity Require Distinct Regions in the SidE Family of Legionella pneumophila Effectors
Abstract
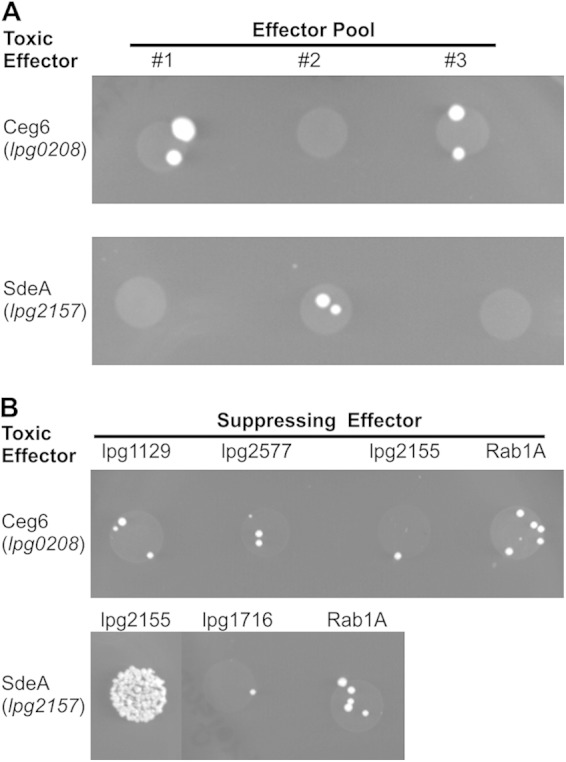
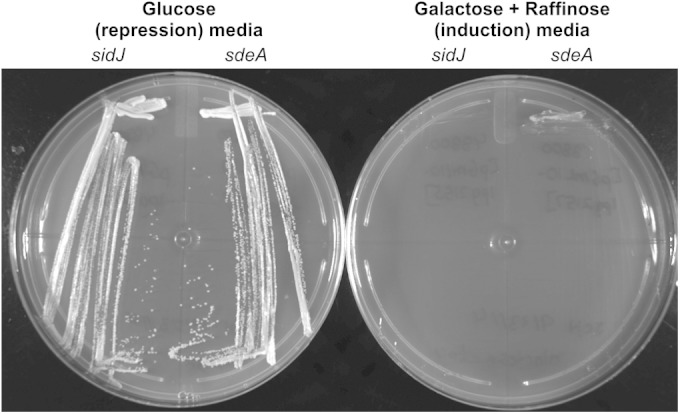
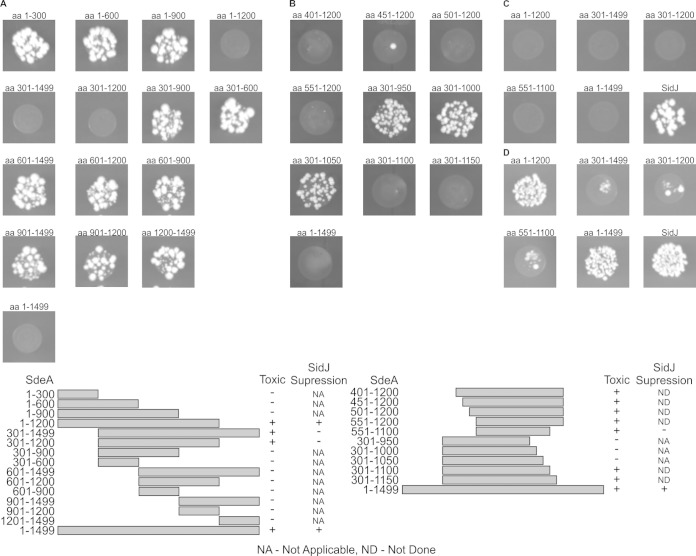
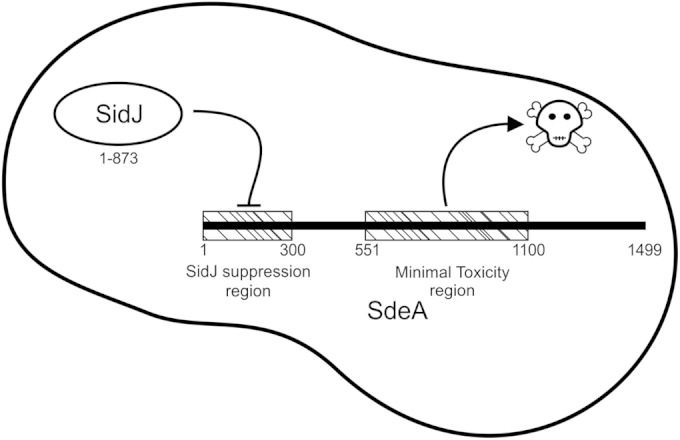
Intracellular bacteria use a variety of strategies to evade degradation and create a replicative niche. Legionella pneumophila is an intravacuolar pathogen that establishes a replicative niche through the secretion of more than 300 effector proteins. The function of most effectors remains to be determined. Toxicity in yeast has been used to identify functional domains and elucidate the biochemical function of effectors. A library of L. pneumophila effectors was screened using an expression plasmid that produces low levels of each protein. This screen identified the effector SdeA as a protein that confers a strong toxic phenotype that inhibits yeast replication. The toxicity of SdeA was suppressed in cells producing the effector SidJ. The effector SdeA is a member of the SidE family of L. pneumophila effector proteins. All SidE orthologs encoded by the Philadelphia isolate of Legionella pneumophila were toxic to yeast, and SidJ suppressed the toxicity of each. We identified a conserved central region in the SidE proteins that was sufficient to mediate yeast toxicity. Surprisingly, SidJ did not suppress toxicity when this central region was produced in yeast. We determined that the amino-terminal region of SidE was essential for SidJ-mediated suppression of toxicity. Thus, there is a genetic interaction that links the activity of SidJ and the amino-terminal region of SidE, which is required to modulate the toxic activity displayed by the central region of the SidE protein. This suggests a complex mechanism by which the L. pneumophila effector SidJ modulates the function of the SidE proteins after translocation into host cells.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
