

DNA polymerase from temperate phage Bam35 is endowed with processive polymerization and abasic sites translesion synthesis capacity
- PMID: 26100910
- PMCID: PMC4500267
- DOI: 10.1073/pnas.1510280112
DNA polymerase from temperate phage Bam35 is endowed with processive polymerization and abasic sites translesion synthesis capacity
Abstract
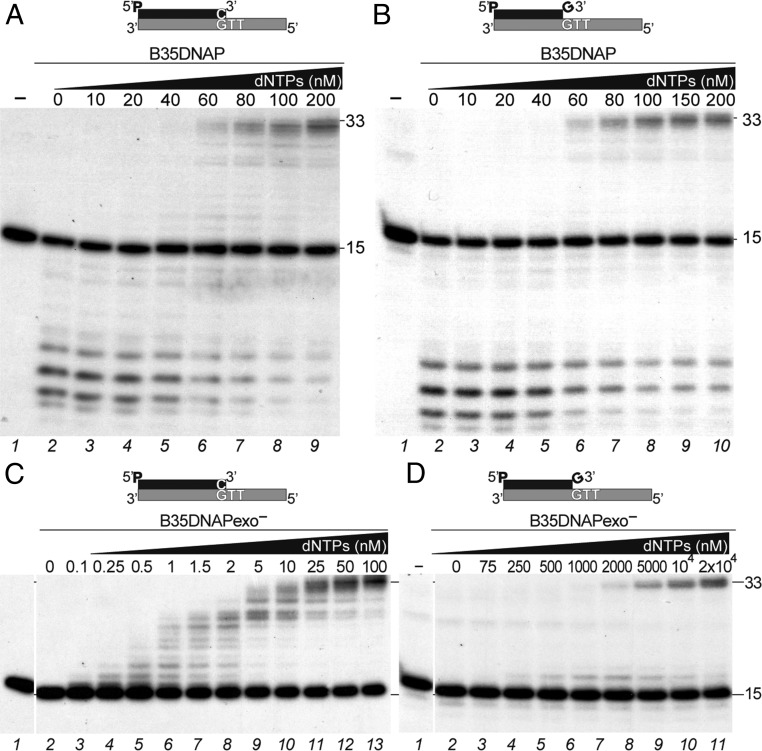
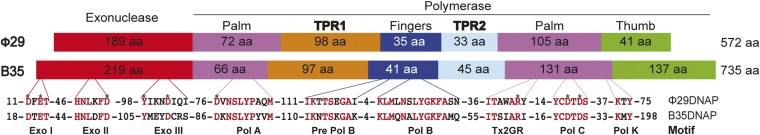
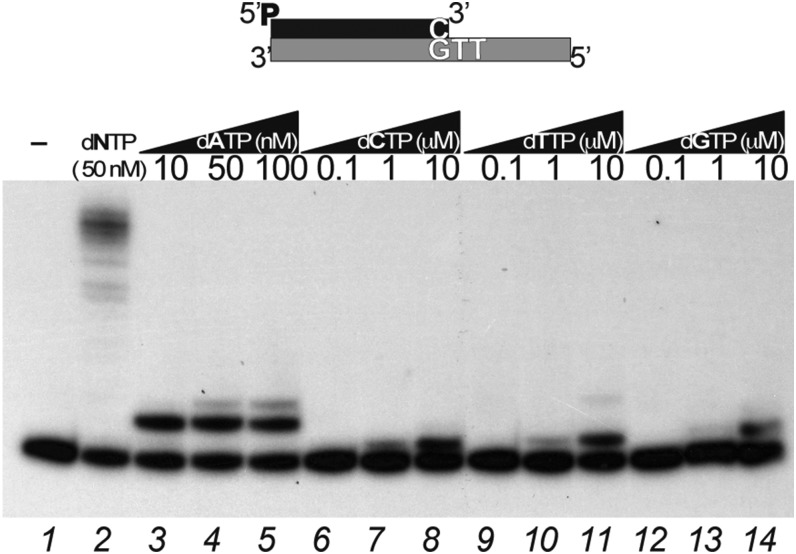
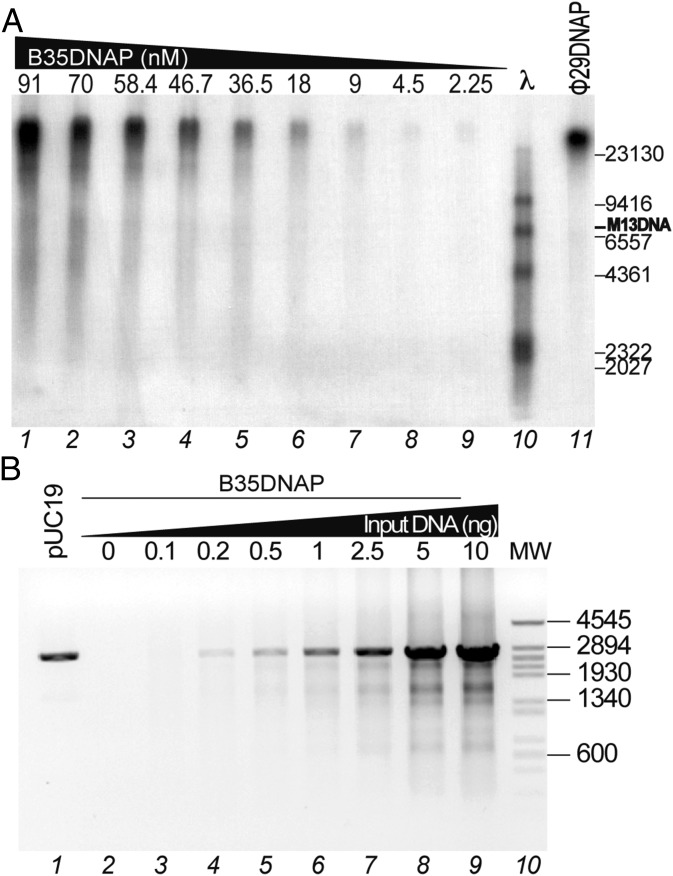
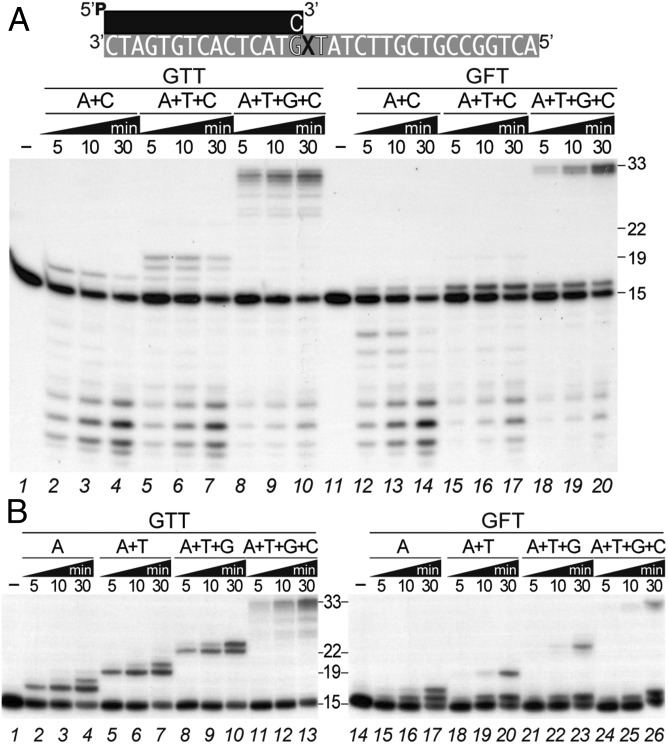
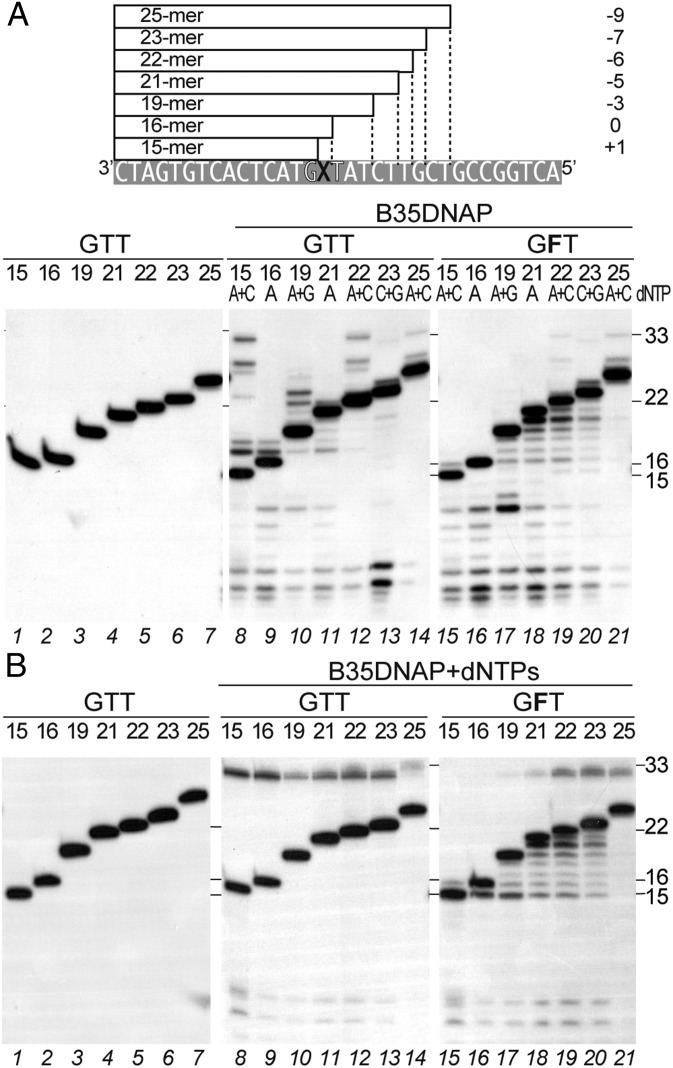
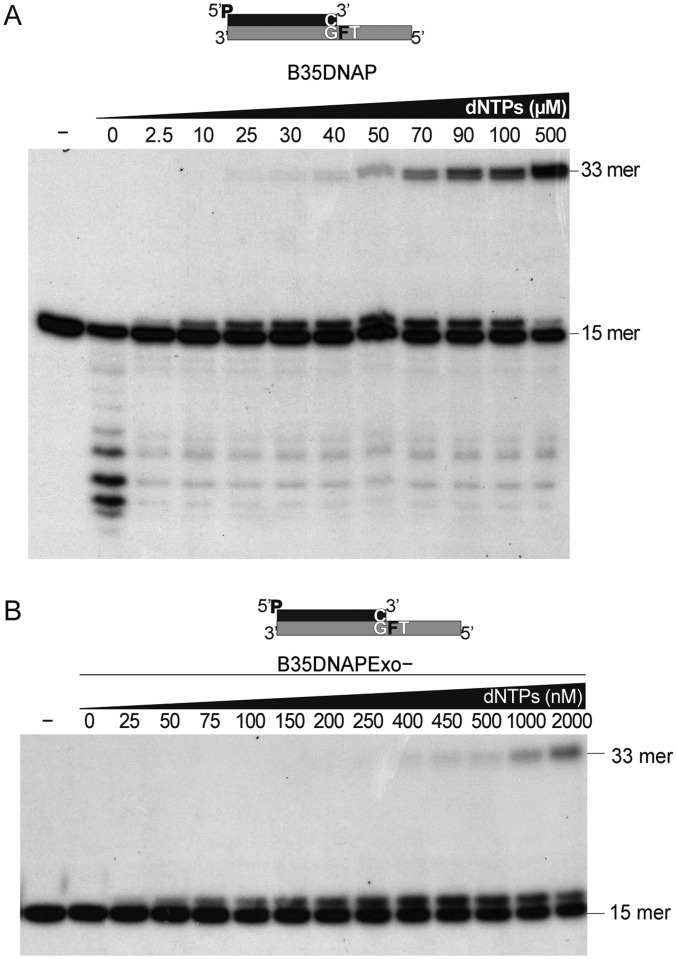
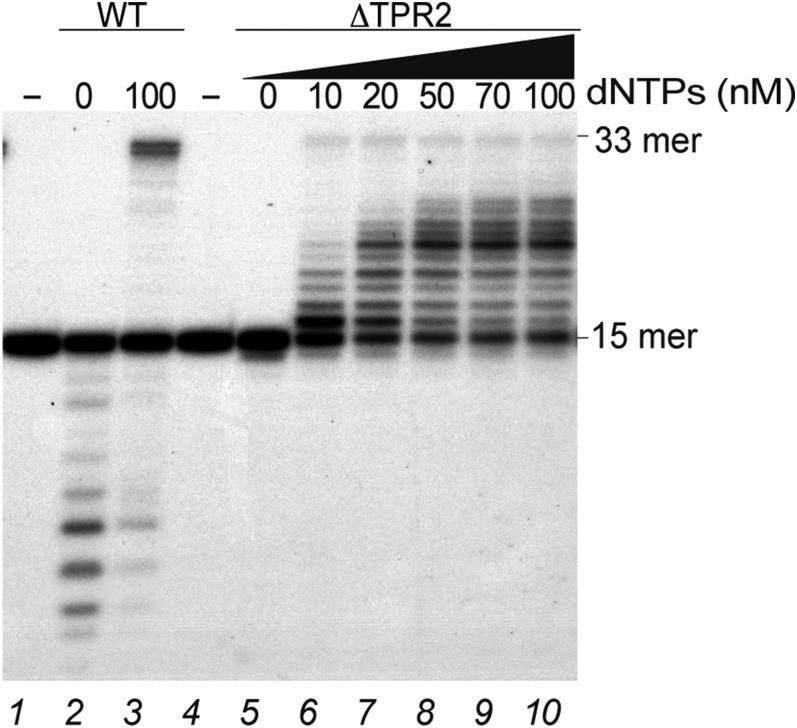
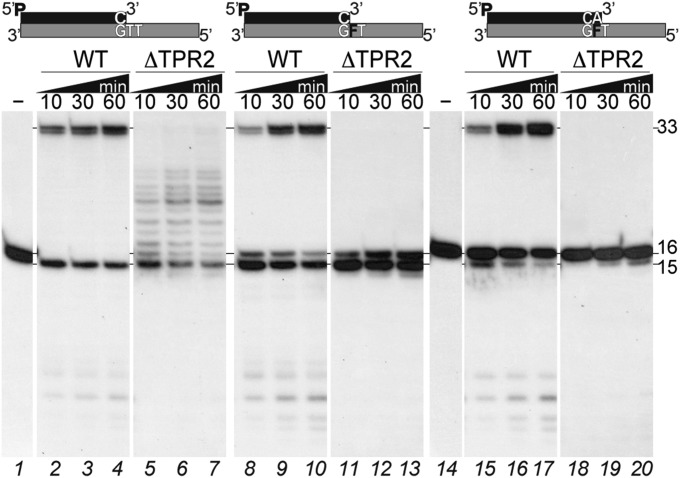
DNA polymerases (DNAPs) responsible for genome replication are highly faithful enzymes that nonetheless cannot deal with damaged DNA. In contrast, translesion synthesis (TLS) DNAPs are suitable for replicating modified template bases, although resulting in very low-fidelity products. Here we report the biochemical characterization of the temperate bacteriophage Bam35 DNA polymerase (B35DNAP), which belongs to the protein-primed subgroup of family B DNAPs, along with phage Φ29 and other viral and mobile element polymerases. B35DNAP is a highly faithful DNAP that can couple strand displacement to processive DNA synthesis. These properties allow it to perform multiple displacement amplification of plasmid DNA with a very low error rate. Despite its fidelity and proofreading activity, B35DNAP was able to successfully perform abasic site TLS without template realignment and inserting preferably an A opposite the abasic site (A rule). Moreover, deletion of the TPR2 subdomain, required for processivity, impaired primer extension beyond the abasic site. Taken together, these findings suggest that B35DNAP may perform faithful and processive genome replication in vivo and, when required, TLS of abasic sites.
Keywords: Bam35; abasic sites; isothermal DNA amplification; protein-primed DNA polymerase; translesion synthesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
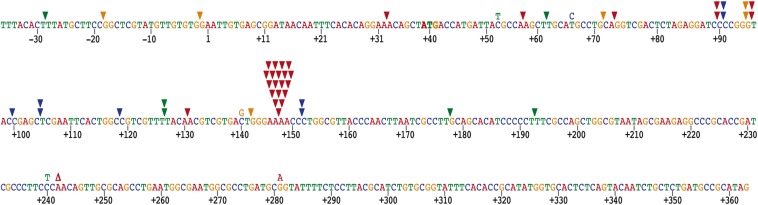
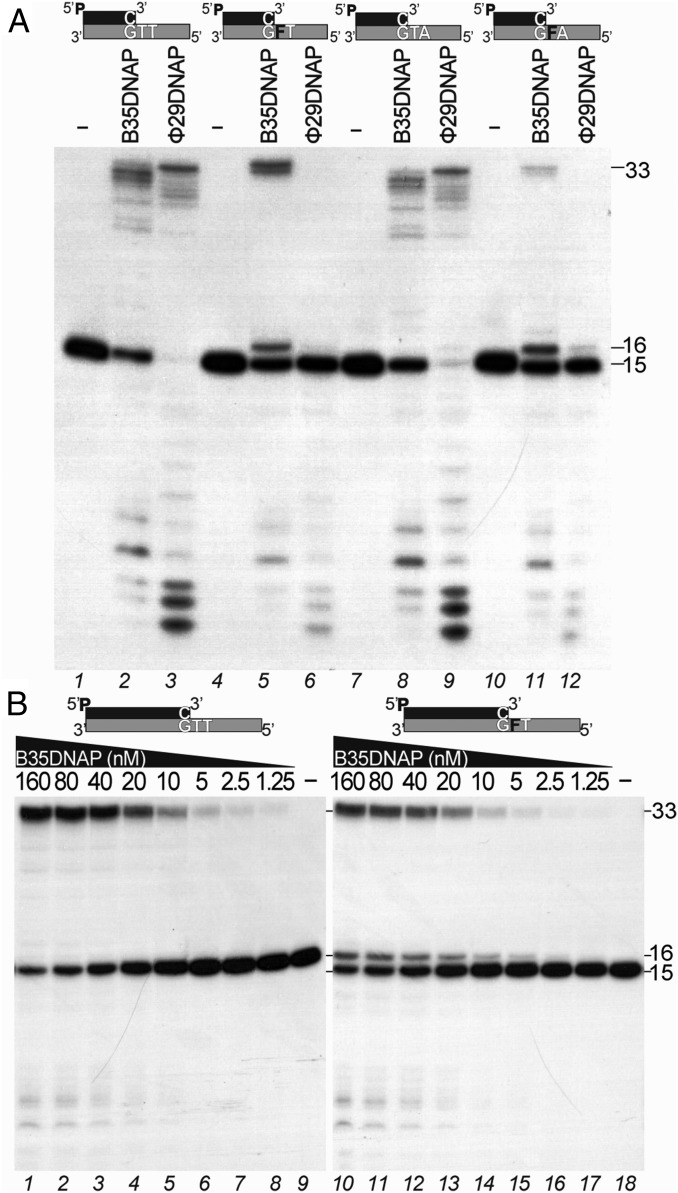
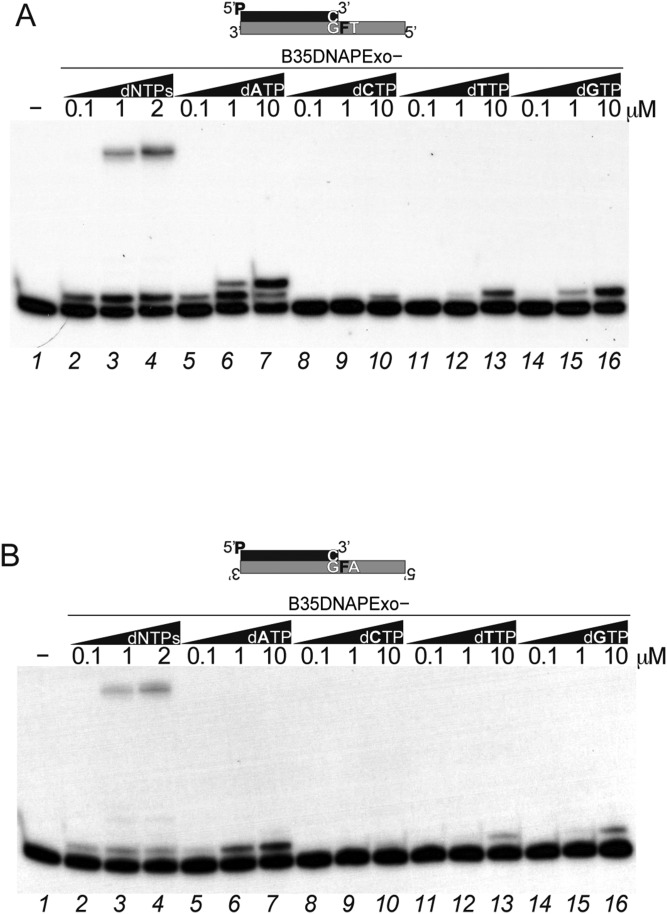
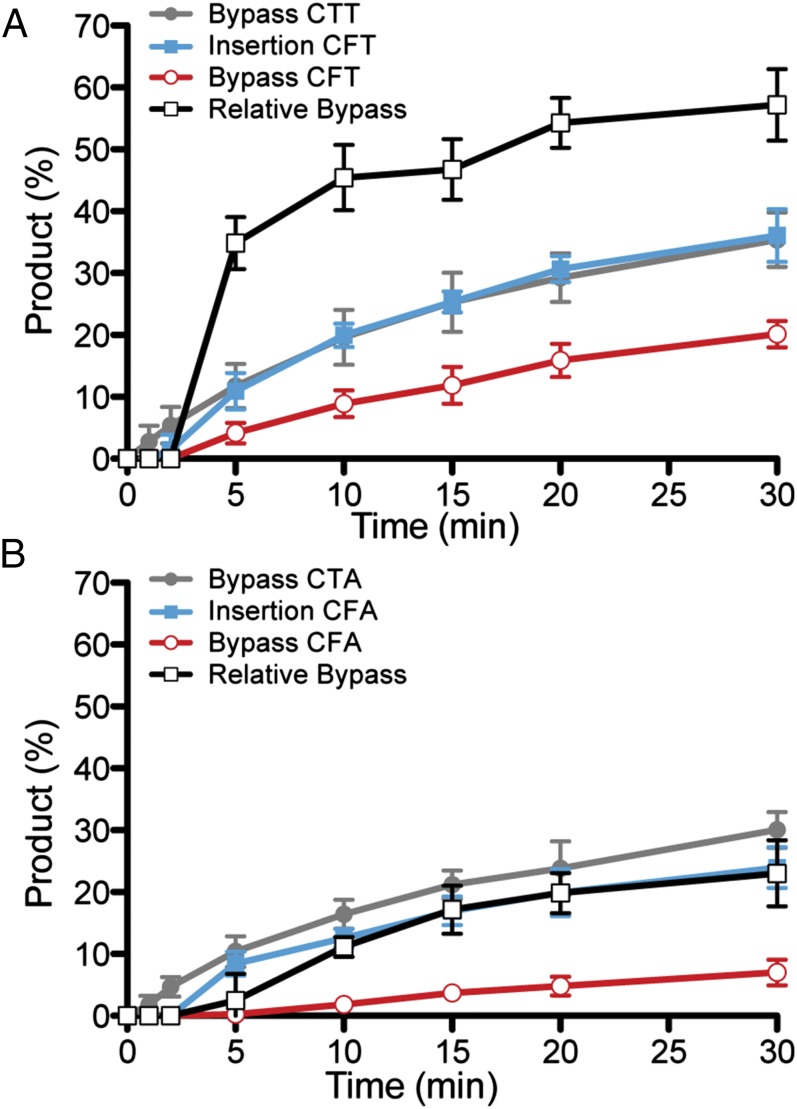
References
-
- Branzei D, Foiani M. Maintaining genome stability at the replication fork. Nat Rev Mol Cell Biol. 2010;11(3):208–219. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
