

Glutamatergic Mechanisms Associated with Seizures and Epilepsy
- PMID: 26101204
- PMCID: PMC4526718
- DOI: 10.1101/cshperspect.a022863
Glutamatergic Mechanisms Associated with Seizures and Epilepsy
Abstract
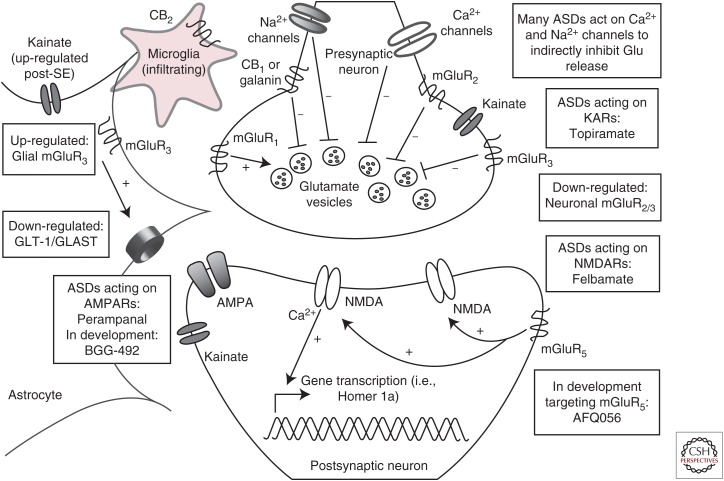
Epilepsy is broadly characterized by aberrant neuronal excitability. Glutamate is the predominant excitatory neurotransmitter in the adult mammalian brain; thus, much of past epilepsy research has attempted to understand the role of glutamate in seizures and epilepsy. Seizures induce elevations in extracellular glutamate, which then contribute to excitotoxic damage. Chronic seizures can alter neuronal and glial expression of glutamate receptors and uptake transporters, further contributing to epileptogenesis. Evidence points to a shared glutamate pathology for epilepsy and other central nervous system (CNS) disorders, including depression, which is often a comorbidity of epilepsy. Therapies that target glutamatergic neurotransmission are available, but many have met with difficulty because of untoward adverse effects. Better understanding of this system has generated novel therapeutic targets that directly and indirectly modulate glutamatergic signaling. Thus, future efforts to manage the epileptic patient with glutamatergic-centric treatments now hold greater potential.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Alexander GM, Godwin DW. 2006. Unique presynaptic and postsynaptic roles of group II metabotropic glutamate receptors in the modulation of thalamic network activity. Neuroscience 141: 501–513. - PubMed
-
- Alexopoulos H, Kosmidis ML, Dalmau J, Dalakas MC. 2011. Paraneoplastic anti-NMDAR encephalitis: Long-term follow-up reveals persistent serum antibodies. J Neurol 258: 1568–1570. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous