

Np9, a cellular protein of retroviral ancestry restricted to human, chimpanzee and gorilla, binds and regulates ubiquitin ligase MDM2
- PMID: 26103464
- PMCID: PMC4614042
- DOI: 10.1080/15384101.2015.1064565
Np9, a cellular protein of retroviral ancestry restricted to human, chimpanzee and gorilla, binds and regulates ubiquitin ligase MDM2
Abstract
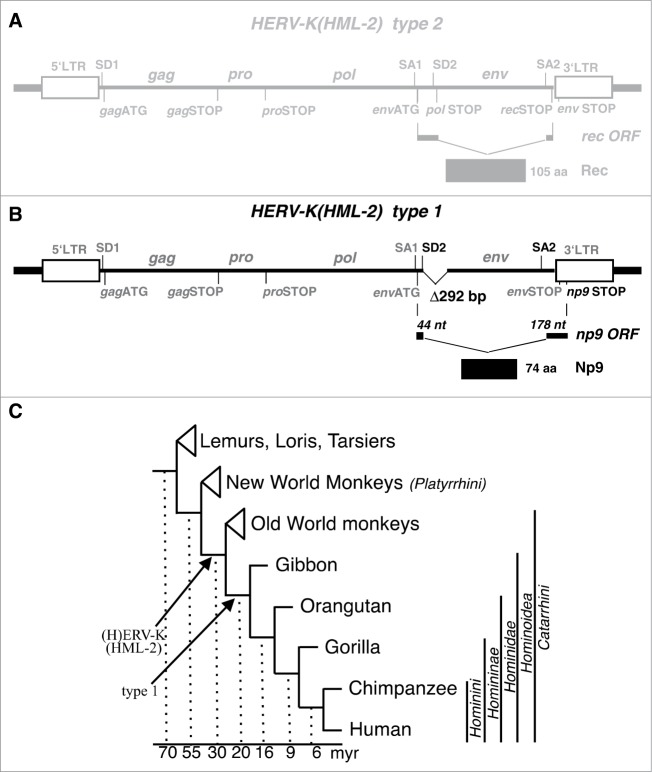
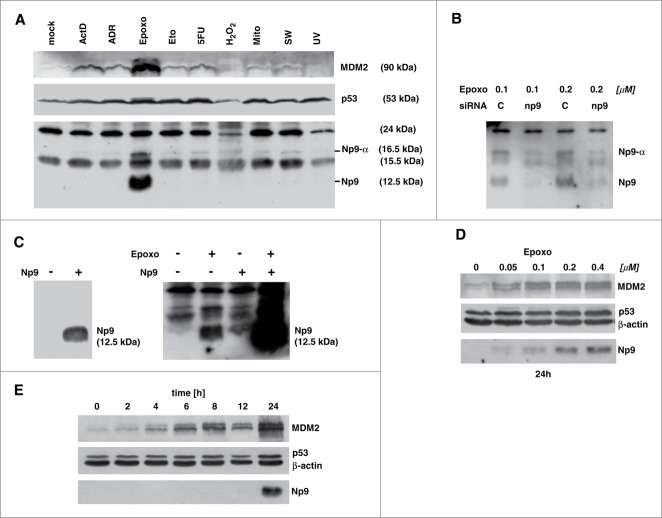
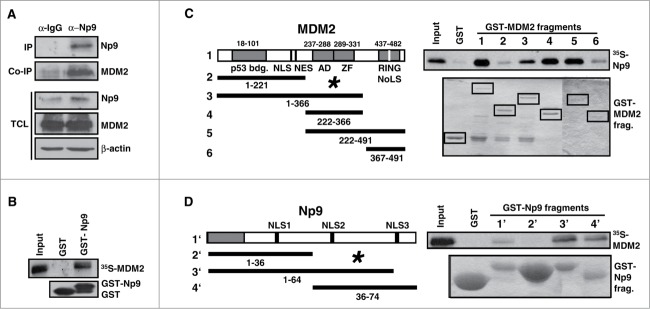
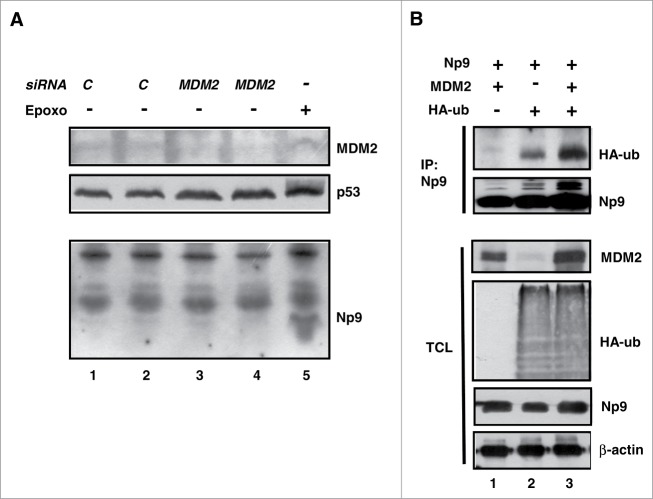
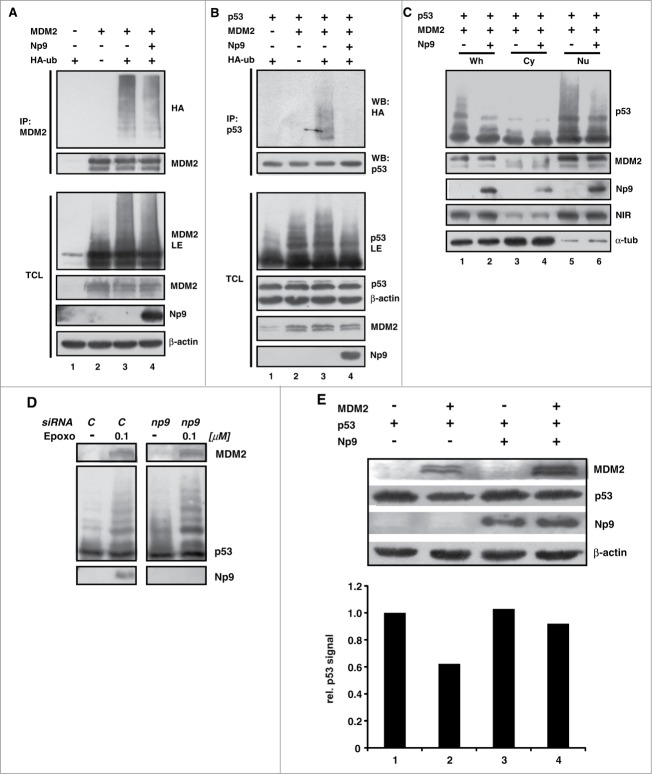
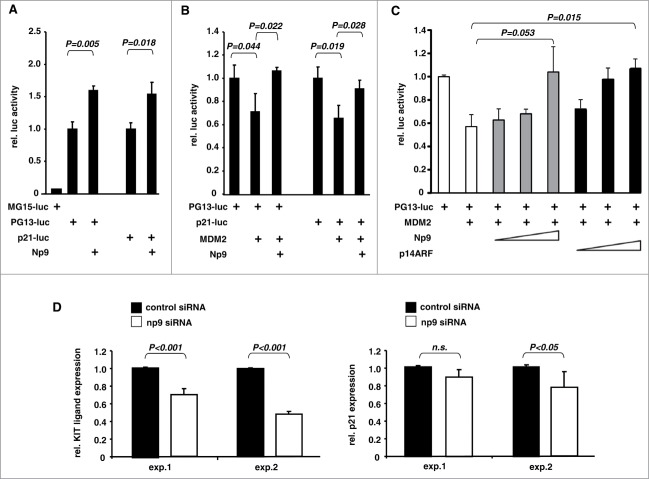
Humans and primates are long-lived animals with long reproductive phases. One factor that appears to contribute to longevity and fertility in humans, as well as to cancer-free survival, is the transcription factor and tumor suppressor p53, controlled by its main negative regulator MDM2. However, p53 and MDM2 homologs are found throughout the metazoan kingdom from Trichoplacidae to Hominidae. Therefore the question arises, if p53/MDM2 contributes to the shaping of primate features, then through which mechanisms. Previous findings have indicated that the appearances of novel p53-regulated genes and wild-type p53 variants during primate evolution are important in this context. Here, we report on another mechanism of potential relevance. Human endogenous retrovirus K subgroup HML-2 (HERV-K(HML-2)) type 1 proviral sequences were formed in the genomes of the predecessors of contemporary Hominoidea and can be identified in the genomes of Nomascus leucogenys (gibbon) up to Homo sapiens. We previously reported on an alternative splicing event in HERV-K(HML-2) type 1 proviruses that can give rise to nuclear protein of 9 kDa (Np9). We document here the evolution of Np9-coding capacity in human, chimpanzee and gorilla, and show that the C-terminal half of Np9 binds directly to MDM2, through a domain of MDM2 that is known to be contacted by various cellular proteins in response to stress. Np9 can inhibit the MDM2 ubiquitin ligase activity toward p53 in the cell nucleus, and can support the transactivation of genes by p53. Our findings point to the possibility that endogenous retrovirus protein Np9 contributes to the regulation of the p53-MDM2 pathway specifically in humans, chimpanzees and gorillas.
Keywords: MDM2; Np9; endogenous retrovirus; evolution; p53; ubiquitylation.
Figures

Similar articles
-
The NP9 protein encoded by the human endogenous retrovirus HERV-K(HML-2) negatively regulates gene activation of the Epstein-Barr virus nuclear antigen 2 (EBNA2).Int J Cancer. 2011 Sep 1;129(5):1105-15. doi: 10.1002/ijc.25760. Epub 2011 Jan 12. Int J Cancer. 2011. PMID: 21710493
-
HERV-K(HML-2) rec and np9 transcripts not restricted to disease but present in many normal human tissues.Mob DNA. 2015 Mar 5;6:4. doi: 10.1186/s13100-015-0035-7. eCollection 2015. Mob DNA. 2015. PMID: 25750667 Free PMC article.
-
Gorillas have been infected with the HERV-K (HML-2) endogenous retrovirus much more recently than humans and chimpanzees.Proc Natl Acad Sci U S A. 2019 Jan 22;116(4):1337-1346. doi: 10.1073/pnas.1814203116. Epub 2019 Jan 4. Proc Natl Acad Sci U S A. 2019. PMID: 30610173 Free PMC article.
-
Dynamics in the p53-Mdm2 ubiquitination pathway.Cell Cycle. 2004 Jul;3(7):895-9. Epub 2004 Jul 2. Cell Cycle. 2004. PMID: 15254415 Review.
-
Regulation of the p14ARF-Mdm2-p53 pathway: an overview in breast cancer.Exp Mol Pathol. 2006 Oct;81(2):115-22. doi: 10.1016/j.yexmp.2006.07.001. Epub 2006 Aug 17. Exp Mol Pathol. 2006. PMID: 16919268 Review.
Cited by
-
Microorganisms as Shapers of Human Civilization, from Pandemics to Even Our Genomes: Villains or Friends? A Historical Approach.Microorganisms. 2021 Dec 6;9(12):2518. doi: 10.3390/microorganisms9122518. Microorganisms. 2021. PMID: 34946123 Free PMC article. Review.
-
Identification of the novel Np17 oncogene in human leukemia.Aging (Albany NY). 2020 Nov 21;12(23):23647-23667. doi: 10.18632/aging.103808. Epub 2020 Nov 21. Aging (Albany NY). 2020. PMID: 33226963 Free PMC article.
-
Prolonged activity of HERV-K(HML2) in Old World Monkeys accounts for recent integrations and novel recombinant variants.Front Microbiol. 2022 Dec 1;13:1040792. doi: 10.3389/fmicb.2022.1040792. eCollection 2022. Front Microbiol. 2022. PMID: 36532485 Free PMC article.
-
The HERV-K accessory protein Np9 controls viability and migration of teratocarcinoma cells.PLoS One. 2019 Feb 28;14(2):e0212970. doi: 10.1371/journal.pone.0212970. eCollection 2019. PLoS One. 2019. PMID: 30818388 Free PMC article.
-
Beyond pathogens: the intriguing genetic legacy of endogenous retroviruses in host physiology.Front Cell Infect Microbiol. 2024 Apr 9;14:1379962. doi: 10.3389/fcimb.2024.1379962. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38655281 Free PMC article. Review.
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, et al.. Initial sequencing and analysis of the human genome. Nature 2001; 409:860-921; PMID:11237011; http://dx.doi.org/10.1038/35057062 - DOI - PubMed
-
- Stoye JP. Studies of endogenous retroviruses reveal a continuing evolutionary saga. Nat Rev Microbiol 2012; 10:395-406; PMID:22565131 - PubMed
-
- Blomberg J, Benachenhou F, Blikstad V, Sperber G, Mayer J. Classification and nomenclature of endogenous retroviral sequences (ERVs): problems and recommendations. Gene 2009; 448:115-23; PMID:19540319; http://dx.doi.org/10.1016/j.gene.2009.06.007 - DOI - PubMed
-
- Mayer J, Blomberg J, Seal RL. A revised nomenclature for transcribed human endogenous retroviral loci. Mob DNA 2011; 2:7; PMID:21542922; http://dx.doi.org/10.1186/1759-8753-2-7 - DOI - PMC - PubMed
-
- Flockerzi A, Ruggieri A, Frank O, Sauter M, Maldener E, Kopper B, Wullich B, Seifarth W, Müller-Lantzsch N, Leib-Mösch C, et al.. Expression patterns of transcribed human endogenous retrovirus HERV-K(HML-2) loci in human tissues and the need for a HERV Transcriptome Project. BMC Genomics 2008; 9:354; PMID:18664271; http://dx.doi.org/10.1186/1471-2164-9-354 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous