

Diversity-generating Retroelements in Phage and Bacterial Genomes
- PMID: 26104433
- PMCID: PMC4498404
- DOI: 10.1128/microbiolspec.MDNA3-0029-2014
Diversity-generating Retroelements in Phage and Bacterial Genomes
Abstract
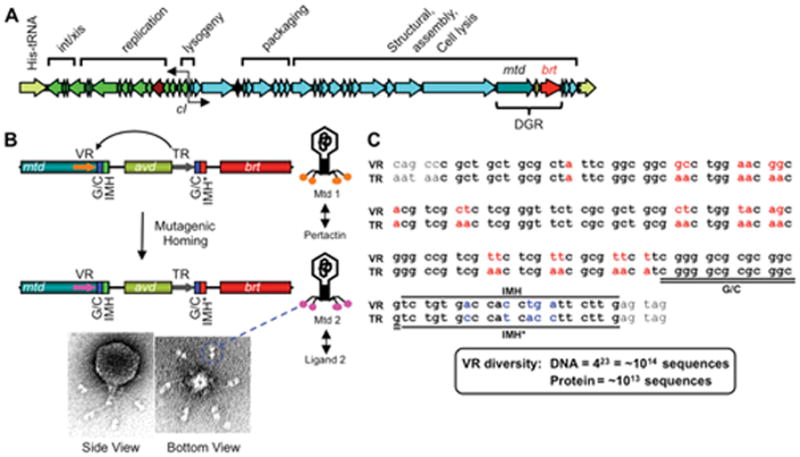
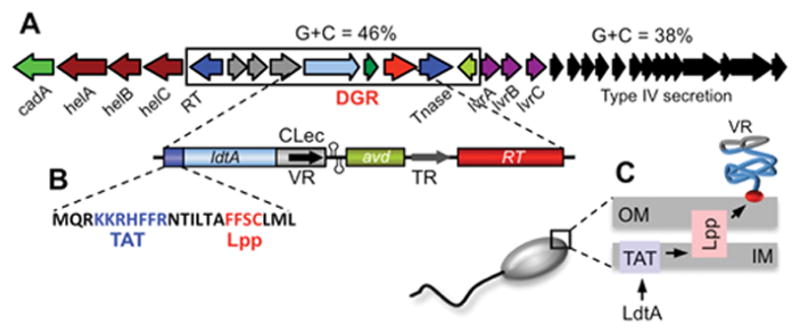
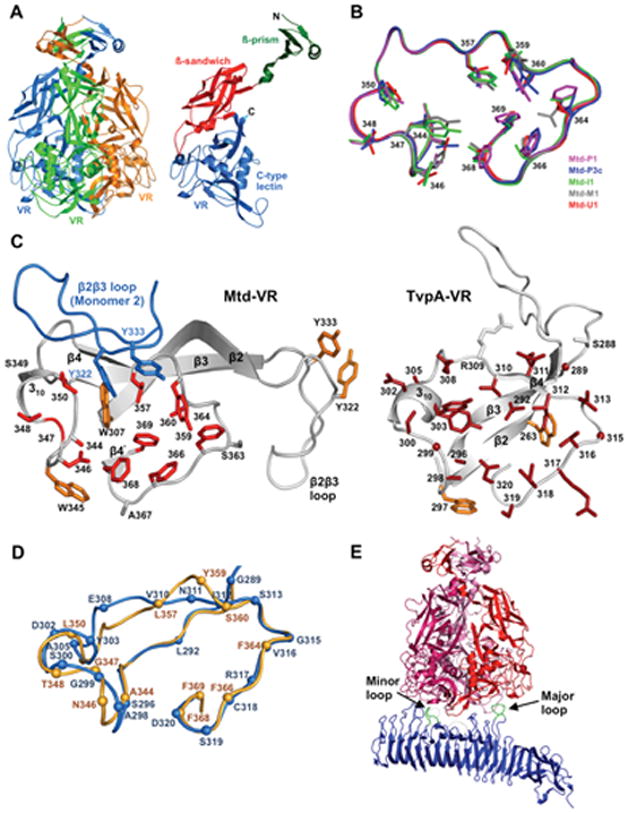
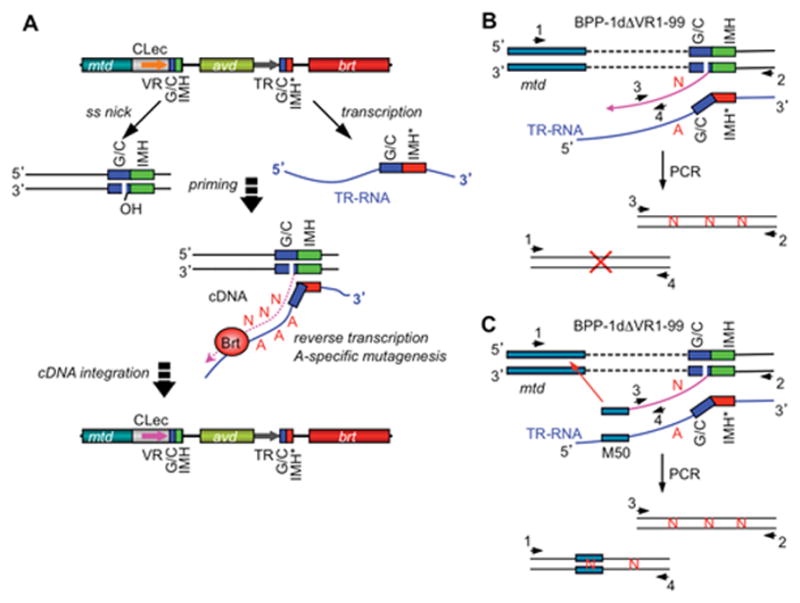
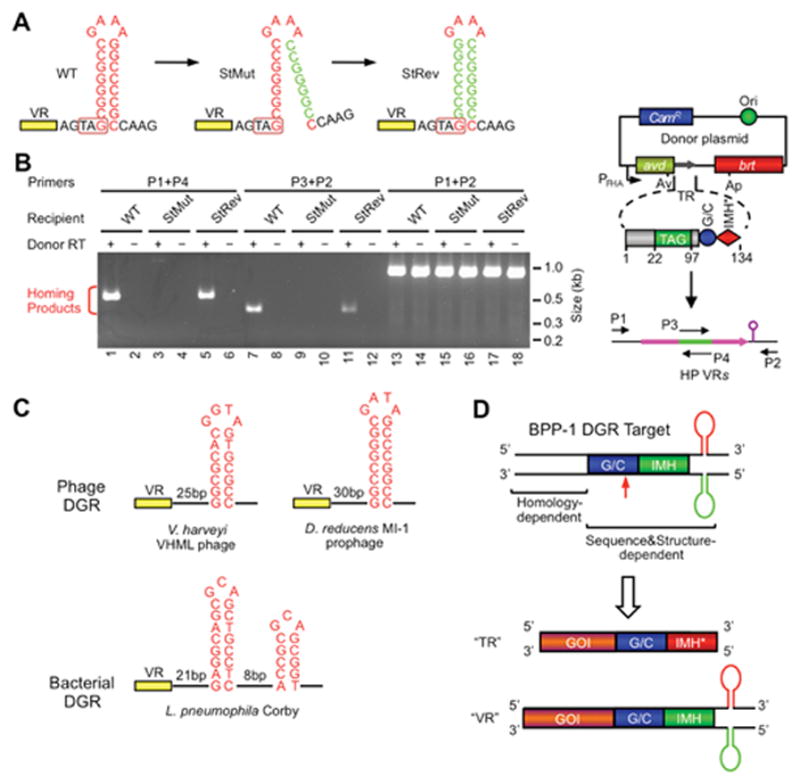
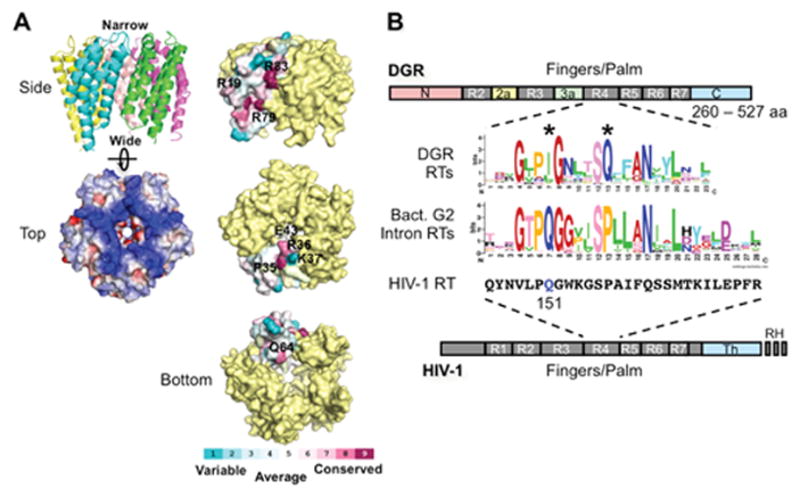
Diversity-generating retroelements (DGRs) are DNA diversification machines found in diverse bacterial and bacteriophage genomes that accelerate the evolution of ligand-receptor interactions. Diversification results from a unidirectional transfer of sequence information from an invariant template repeat (TR) to a variable repeat (VR) located in a protein-encoding gene. Information transfer is coupled to site-specific mutagenesis in a process called mutagenic homing, which occurs through an RNA intermediate and is catalyzed by a unique, DGR-encoded reverse transcriptase that converts adenine residues in the TR into random nucleotides in the VR. In the prototype DGR found in the Bordetella bacteriophage BPP-1, the variable protein Mtd is responsible for phage receptor recognition. VR diversification enables progeny phage to switch tropism, accelerating their adaptation to changes in sequence or availability of host cell-surface molecules for infection. Since their discovery, hundreds of DGRs have been identified, and their functions are just beginning to be understood. VR-encoded residues of many DGR-diversified proteins are displayed in the context of a C-type lectin fold, although other scaffolds, including the immunoglobulin fold, may also be used. DGR homing is postulated to occur through a specialized target DNA-primed reverse transcription mechanism that allows repeated rounds of diversification and selection, and the ability to engineer DGRs to target heterologous genes suggests applications for bioengineering. This chapter provides a comprehensive review of our current understanding of this newly discovered family of beneficial retroelements.
Figures
References
-
- Agrawal A, Eastman QM, Schatz DG. Transposition mediated by RAG1 and RAG2 and its implications for the evolution of the immune system. Nature. 1998;394:744–751. - PubMed
-
- Liu M, Deora R, Doulatov SR, Gingery M, Eiserling FA, Preston A, Maskell DJ, Simons RW, Cotter PA, Parkhill J, Miller JF. Reverse transcriptase-mediated tropism switching in Bordetella bacteriophage. Science. 2002;295:2091–2094. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
