

Effects of Ixeris dentata water extract and caffeic acid on allergic inflammation in vivo and in vitro
- PMID: 26104582
- PMCID: PMC4479232
- DOI: 10.1186/s12906-015-0700-x
Effects of Ixeris dentata water extract and caffeic acid on allergic inflammation in vivo and in vitro
Abstract
Background: Ixeris dentata Nakai has been used for the treatment of mithridatism, calculous, indigestion, pneumonia, hepatitis, and tumors in Korea, China, and Japan. However, the effect of a water extract of Ixeris dentata (ID) and its molecular mechanism on allergic inflammation has not been elucidated. In this study, we attempted to evaluate the effects of ID and its major compound caffeic acid on allergic inflammation in vivo and in vitro.
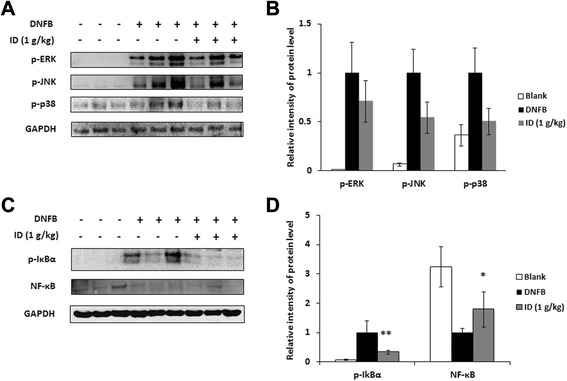
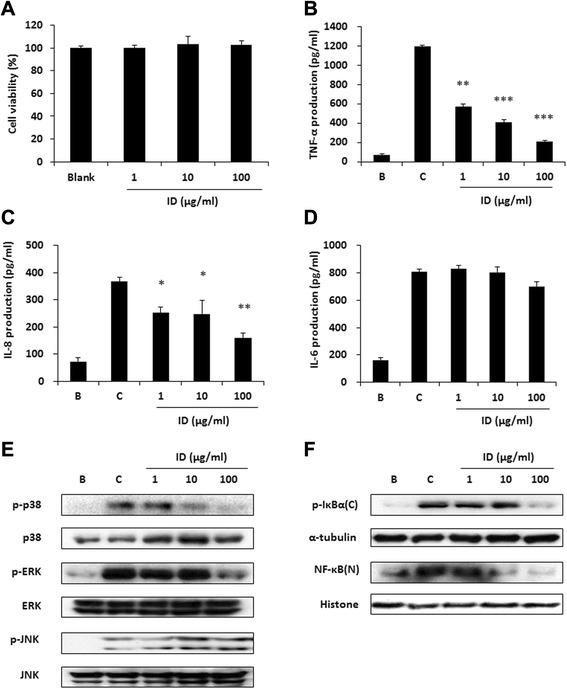
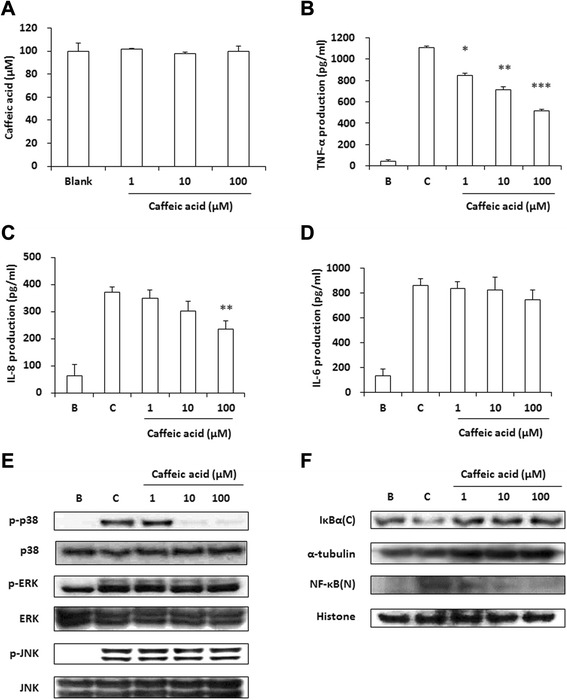
Methods: ID was applied to 2, 4-dinitrofluorobenzene (DNFB)-induced atopic dermatitis (AD)-like skin lesion mice and immune cell infiltration, cytokine production, and the activation of mitogen-activated protein kinases (MAPKs) were investigated. Moreover, the effect of ID on compound 48/80-induced anaphylactic shock was investigated in a mouse model. The human keratinocyte cell line (HaCaT cells) and human mast cells (HMC-1) were treated with ID or caffeic acid to investigate the effects on the production of chemokines and proinflammatory cytokines and on the activation of MAPKs.
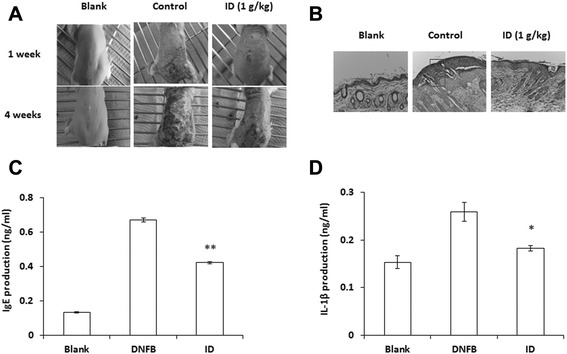
Results: ID inhibited the serum levels of IgE and interleukin (IL)-1β in DNFB-induced AD-like skin lesion mouse models and suppressed anaphylactic shock in the mouse models. ID and caffeic acid inhibited the production of chemokines and adhesion molecules in HaCaT cells. In addition, ID reduced the release of tumor necrosis factor-α and IL-8 via the inhibition of MAPKs phosphorylation in HMC-1 cells.
Conclusions: These results suggest that ID is a potential therapeutic agent for allergic inflammatory diseases, including dermatitis.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
