

LMO2 at 25 years: a paradigm of chromosomal translocation proteins
- PMID: 26108219
- PMCID: PMC4632508
- DOI: 10.1098/rsob.150062
LMO2 at 25 years: a paradigm of chromosomal translocation proteins
Abstract
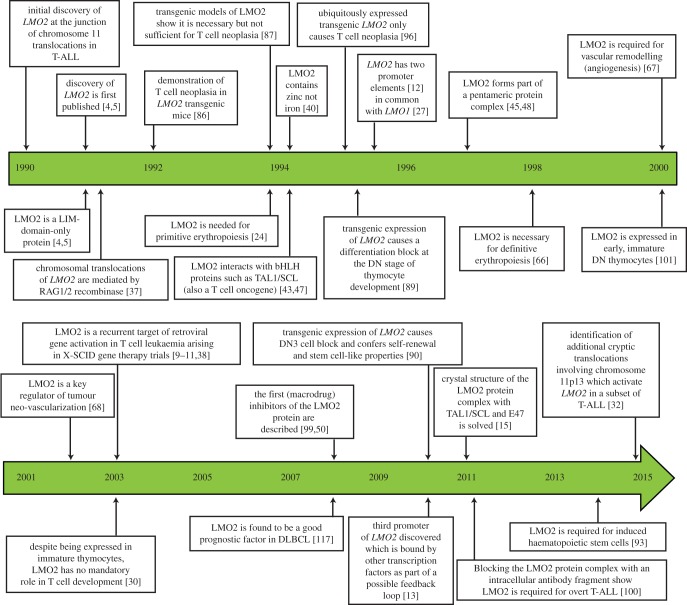
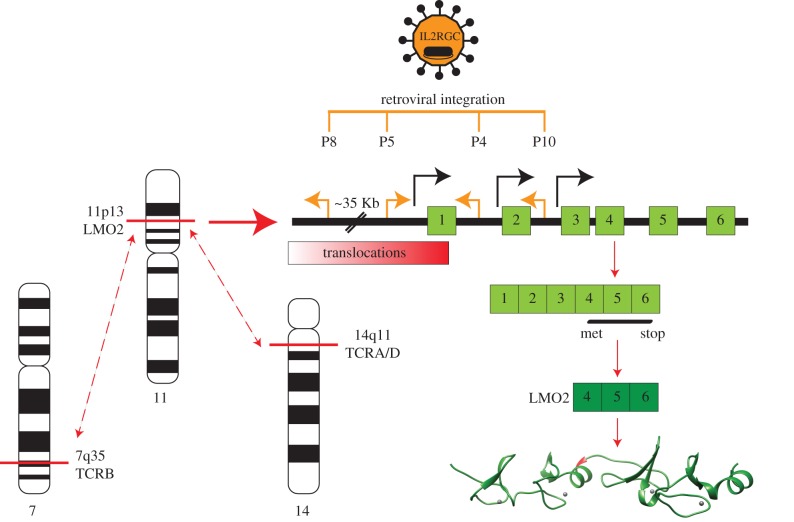
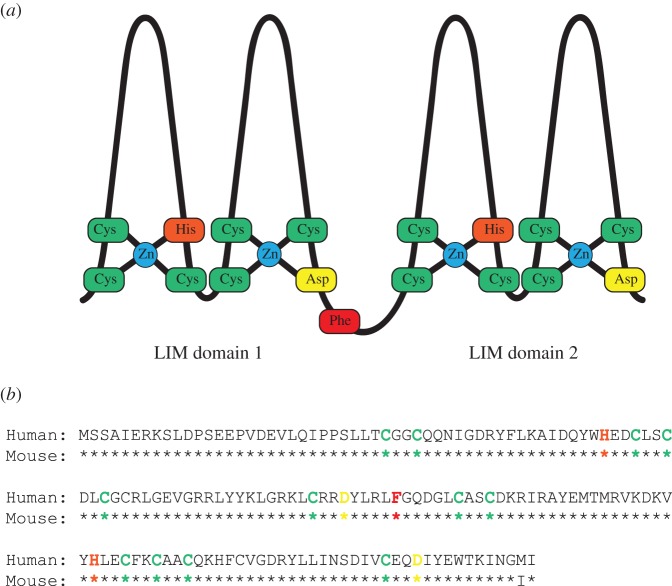
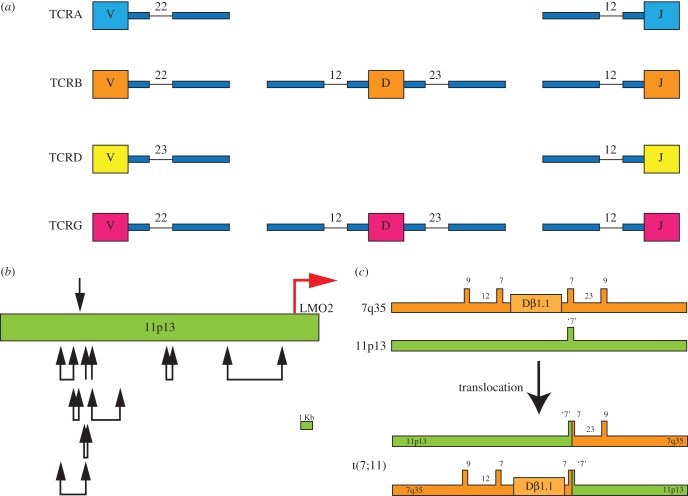
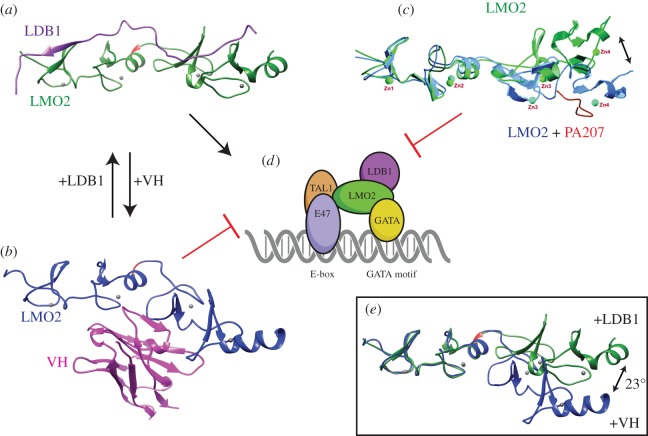
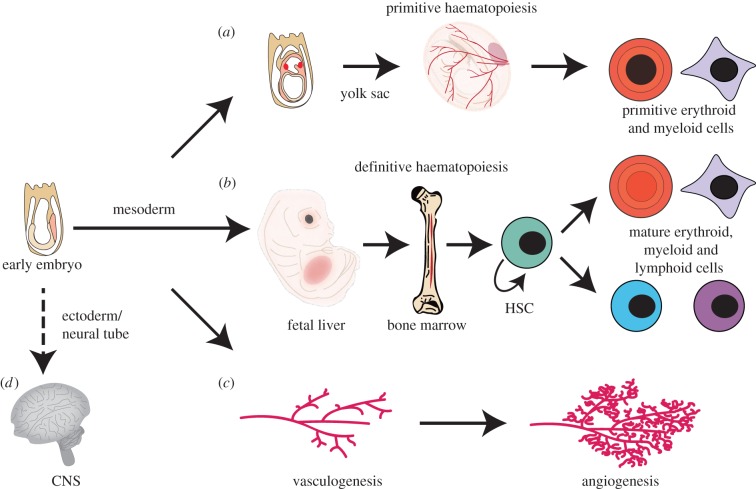
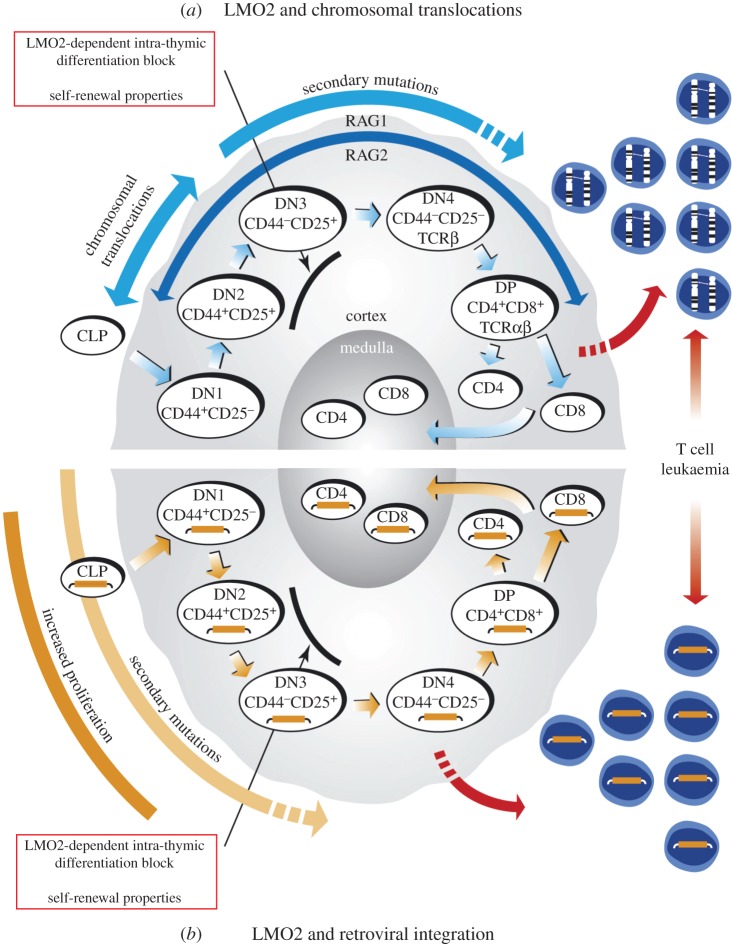
LMO2 was first discovered through proximity to frequently occurring chromosomal translocations in T cell acute lymphoblastic leukaemia (T-ALL). Subsequent studies on its role in tumours and in normal settings have highlighted LMO2 as an archetypical chromosomal translocation oncogene, activated by association with antigen receptor gene loci and a paradigm for translocation gene activation in T-ALL. The normal function of LMO2 in haematopoietic cell fate and angiogenesis suggests it is a master gene regulator exerting a dysfunctional control on differentiation following chromosomal translocations. Its importance in T cell neoplasia has been further emphasized by the recurrent findings of interstitial deletions of chromosome 11 near LMO2 and of LMO2 as a target of retroviral insertion gene activation during gene therapy trials for X chromosome-linked severe combined immuno-deficiency syndrome, both types of event leading to similar T cell leukaemia. The discovery of LMO2 in some B cell neoplasias and in some epithelial cancers suggests a more ubiquitous function as an oncogenic protein, and that the current development of novel inhibitors will be of great value in future cancer treatment. Further, the role of LMO2 in angiogenesis and in haematopoietic stem cells (HSCs) bodes well for targeting LMO2 in angiogenic disorders and in generating autologous induced HSCs for application in various clinical indications.
Keywords: LMO2; X chromosome-linked severe combined immuno-deficiency syndrome; angiogenesis; chromosomal translocations; haematopoiesis; leukaemia.
Figures
References
-
- Rabbitts TH. 1991. Translocations, master genes, and differences between the origins of acute and chronic leukemias. Cell 67, 641–644. (doi:10.1016/0092-8674(91)90057-6) - DOI - PubMed
-
- Rabbitts TH, Boehm T. 1991. Structural and functional chimerism results from chromosomal translocation in lymphoid tumors. Adv. Immunol. 50, 119–146. (doi:10.1016/s0065-2776(08)60824-x) - DOI - PubMed
-
- Lefranc MP, Rabbitts TH. 1985. Two tandemly organized human genes encoding the T-cell gamma constant-region sequences show multiple rearrangement in different T-cell types. Nature 316, 464–466. (doi:10.1038/316464a0) - DOI - PubMed
-
- Boehm T, Foroni L, Kaneko Y, Perutz MF, Rabbitts TH. 1991. The rhombotin family of cysteine-rich LIM-domain oncogenes: distinct members are involved in T-cell translocations to human chromosomes 11p15 and 11p13. Proc. Natl Acad. Sci. USA 88, 4367–4371. (doi:10.1073/pnas.88.10.4367) - DOI - PMC - PubMed
-
- Royer-Pokora B, Loos U, Ludwig WD. 1991. TTG-2, a new gene encoding a cysteine-rich protein with the LIM motif, is overexpressed in acute T-cell leukaemia with the t(11;14)(p13;q11). Oncogene 6, 1887–1893. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources