

The role of juvenile hormone and insulin/TOR signaling in the growth of Manduca sexta
- PMID: 26108483
- PMCID: PMC4499214
- DOI: 10.1186/s12915-015-0155-z
The role of juvenile hormone and insulin/TOR signaling in the growth of Manduca sexta
Abstract
Background: In many insect species, fitness trade-offs exist between maximizing body size and developmental speed. Understanding how various species evolve different life history strategies requires knowledge of the physiological mechanisms underlying the regulation of body size and developmental timing. Here the roles of juvenile hormone (JH) and insulin/target of rapamycin (TOR) signaling in the regulation of the final body size were examined in the tobacco hornworm, Manduca sexta.
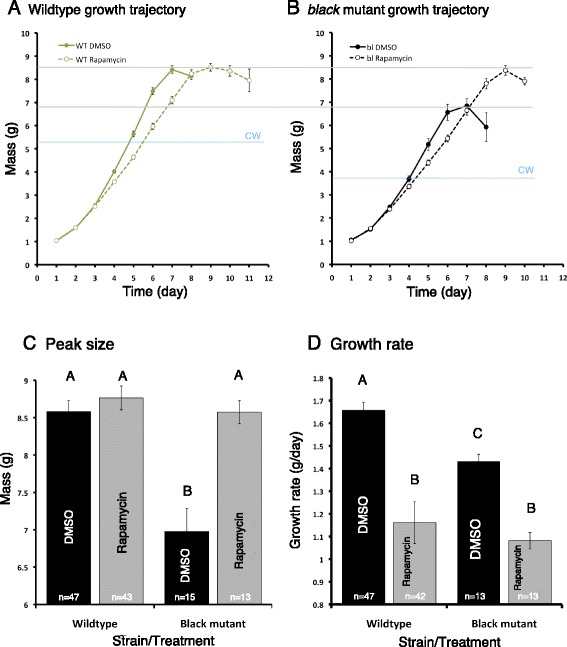
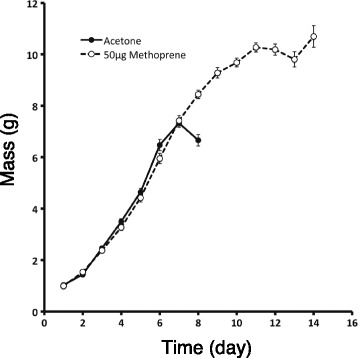
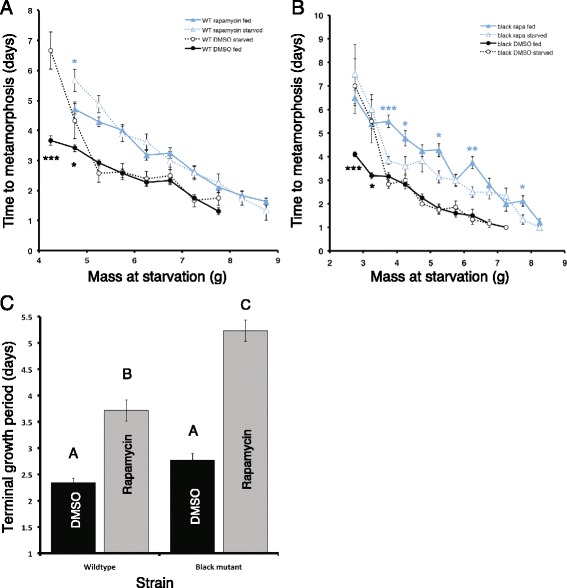
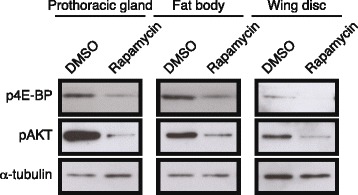
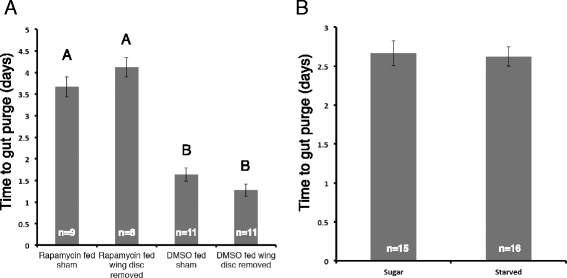
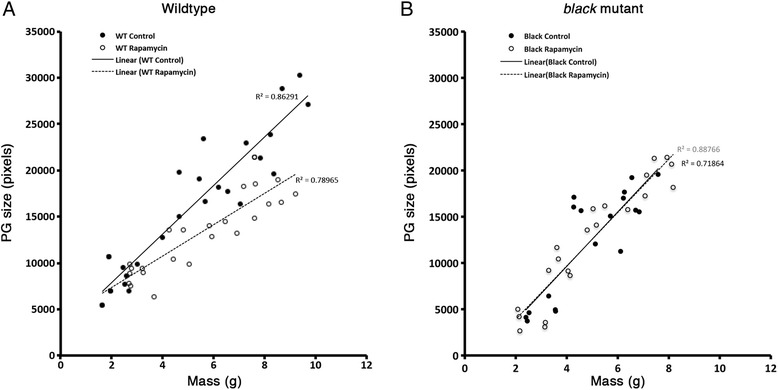
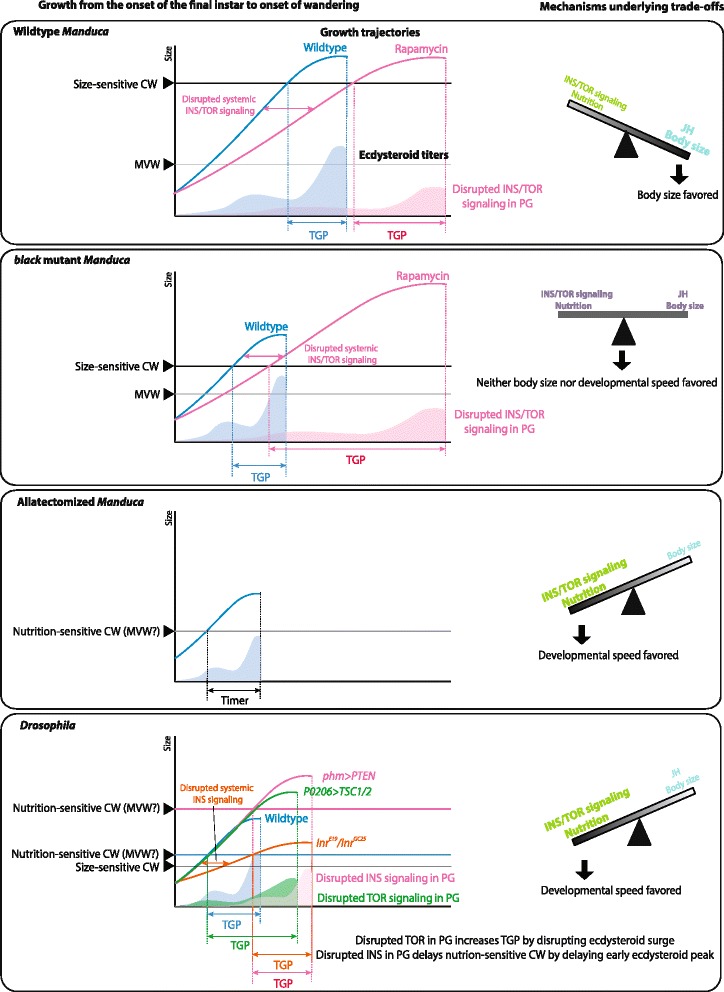
Results: Feeding rapamycin to wild-type larvae decreased the growth rate but did not alter the peak size of the larvae. In contrast, feeding rapamycin to the JH-deficient black mutant larvae caused the larvae to significantly increase the peak size relative to the DMSO-fed control animals by lengthening the terminal growth period. Furthermore, the critical weight was unaltered by feeding rapamycin, indicating that in Manduca, the critical weight is not influenced by insulin/TOR signaling. In addition, post-critical weight starved black mutant Manduca given rapamycin underwent metamorphosis sooner than those that were fed, mimicking the "bail-out mechanism".
Conclusions: Our study demonstrates that JH masks the effects of insulin/TOR signaling in the determination of the final body size and that the critical weights in Drosophila and Manduca rely on distinct mechanisms that reflect different life history strategies. Our study also suggests that TOR signaling lengthens the terminal growth period in Manduca as it does in Drosophila, and that JH levels determine the relative contributions of nutrient- and body size-sensing pathways to metamorphic timing.
Figures
Comment in
-
Big or fast: two strategies in the developmental control of body size.BMC Biol. 2015 Aug 4;13:57. doi: 10.1186/s12915-015-0173-x. BMC Biol. 2015. PMID: 26239356 Free PMC article.
References
-
- Nijhout HF, Williams CM. Control of molting and metamorphosis in tobacco hornworm, Manduca-sexta (L) - cessation of juvenile-hormone secretion as a trigger for pupation. J Exp Biol. 1974;61:493–501. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
