

AKTIP/Ft1, a New Shelterin-Interacting Factor Required for Telomere Maintenance
- PMID: 26110528
- PMCID: PMC4481533
- DOI: 10.1371/journal.pgen.1005167
AKTIP/Ft1, a New Shelterin-Interacting Factor Required for Telomere Maintenance
Abstract
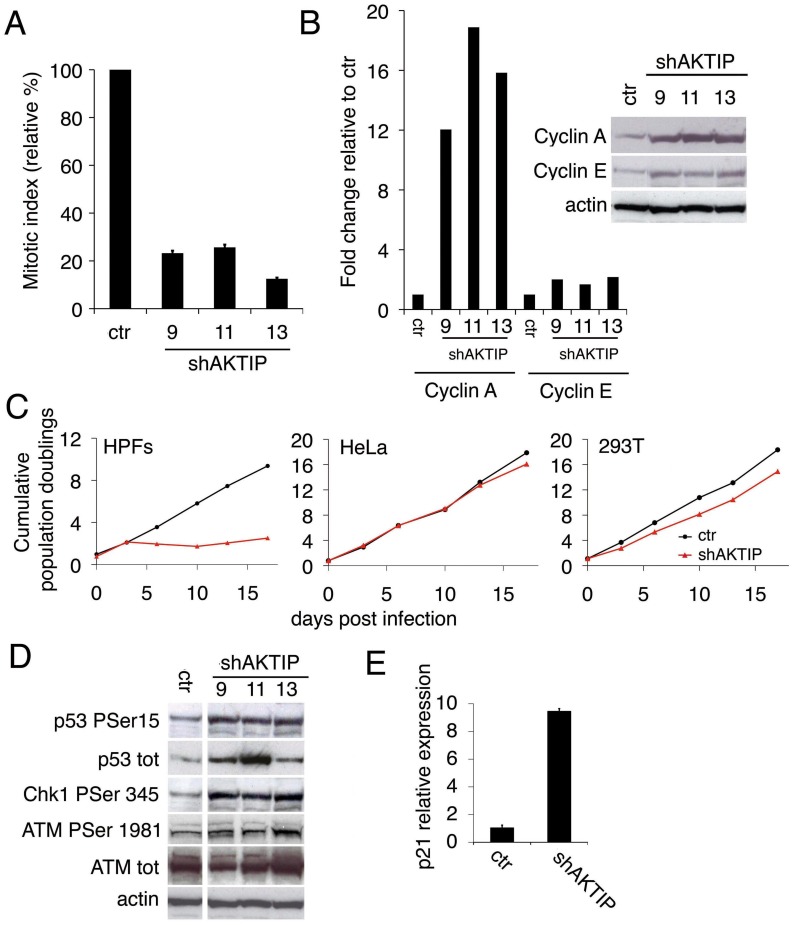
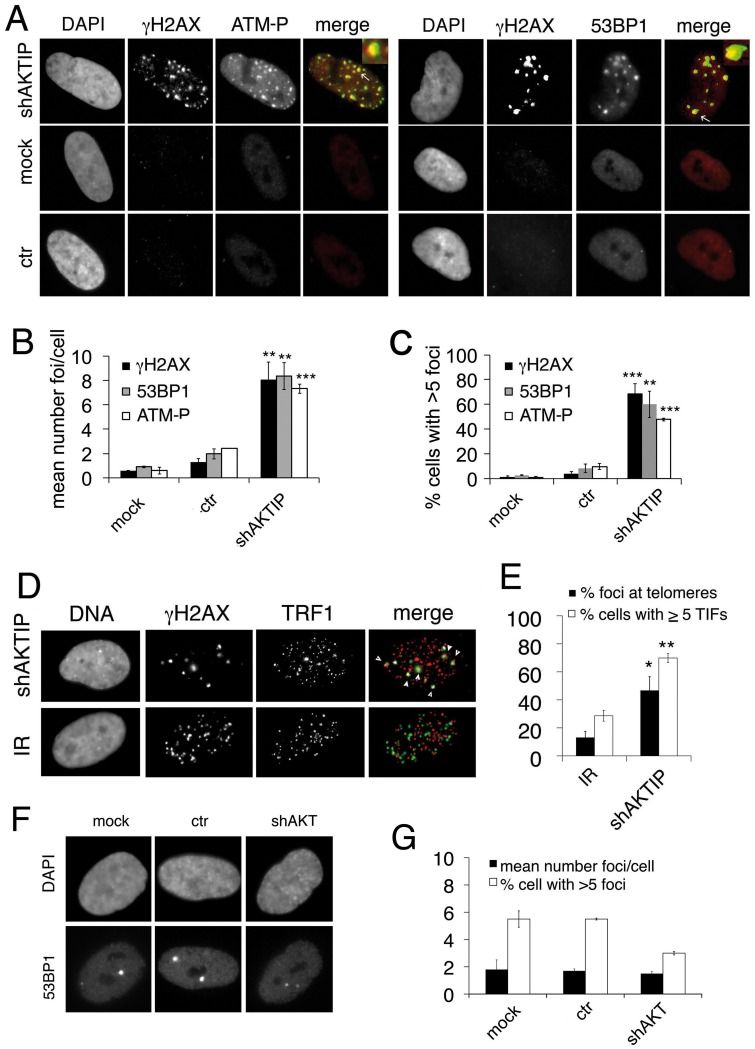
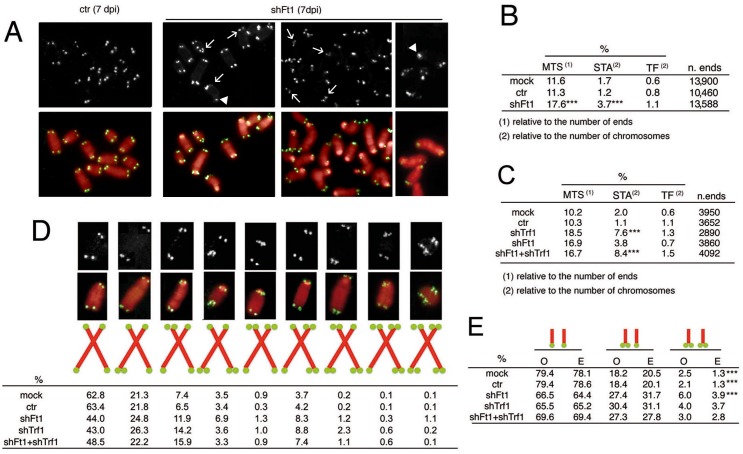
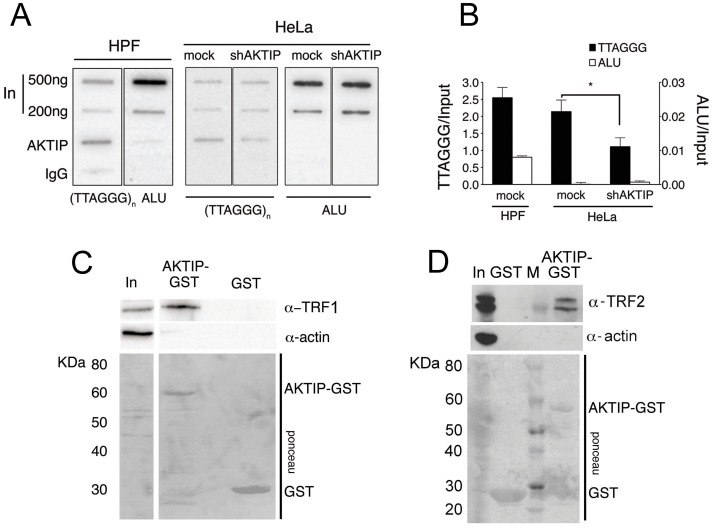
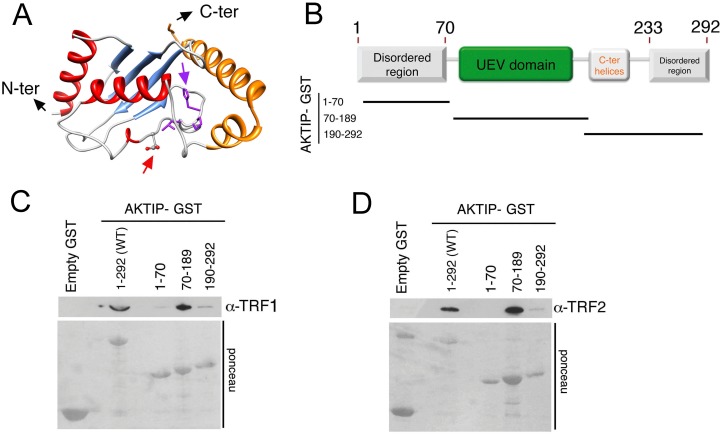
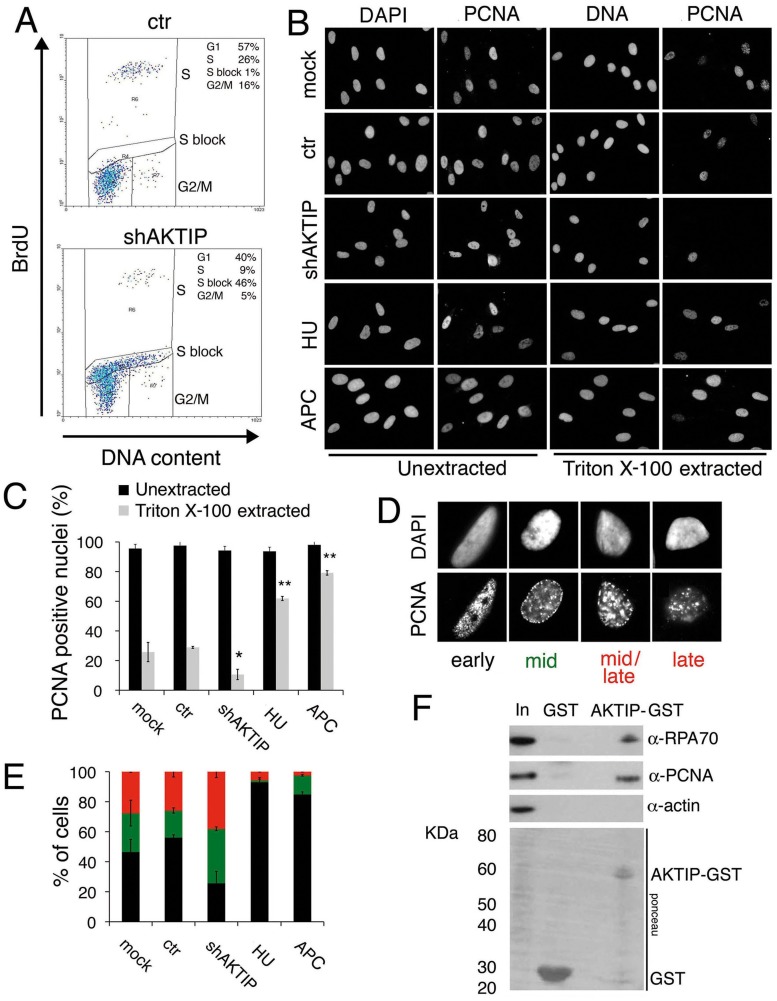
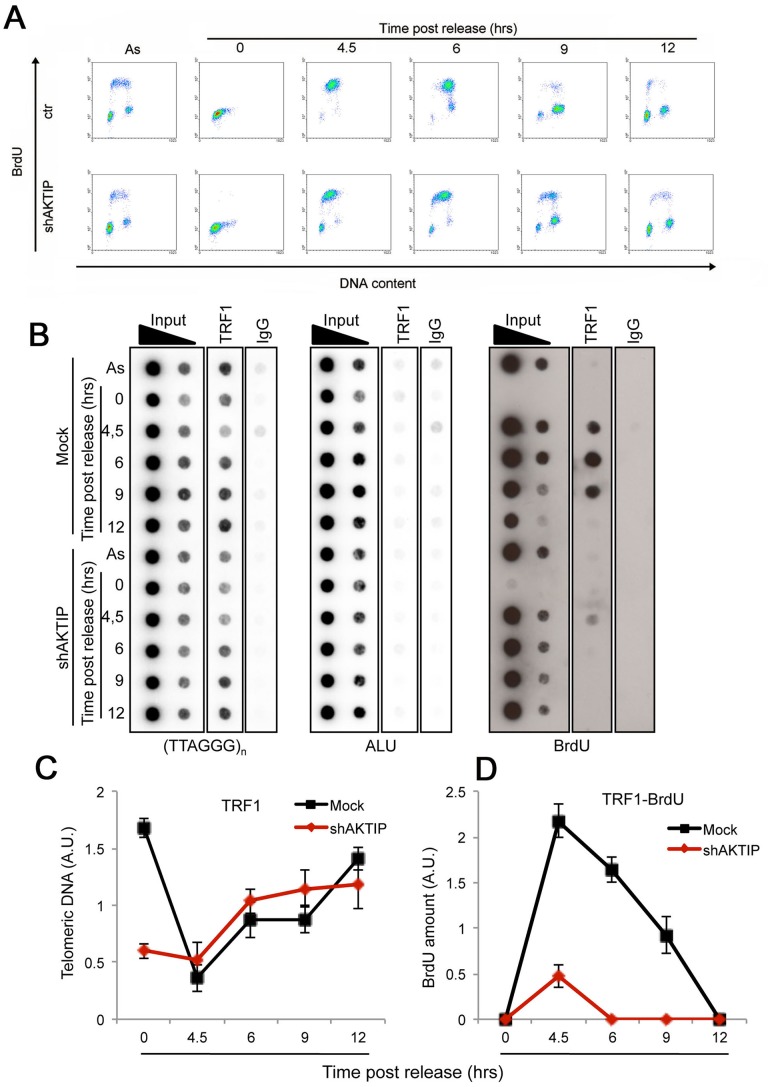
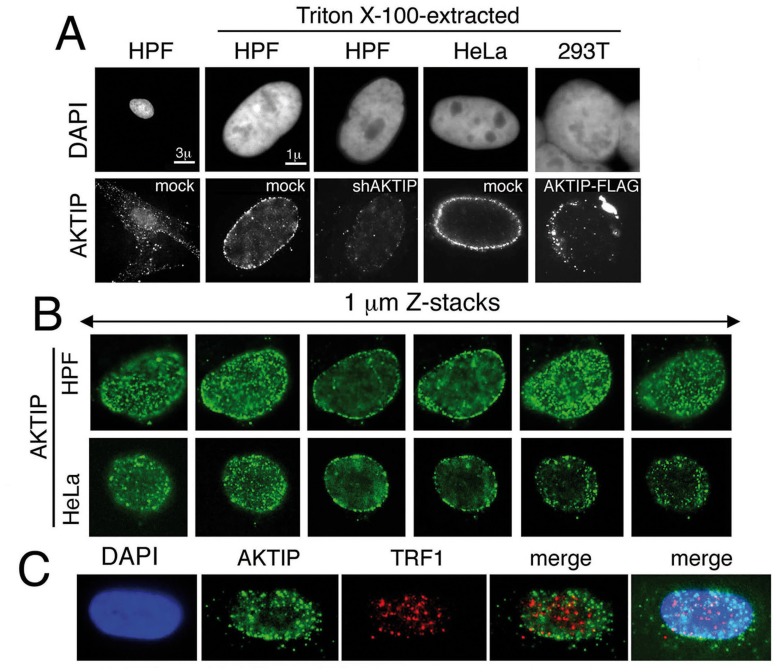
Telomeres are nucleoprotein complexes that protect the ends of linear chromosomes from incomplete replication, degradation and detection as DNA breaks. Mammalian telomeres are protected by shelterin, a multiprotein complex that binds the TTAGGG telomeric repeats and recruits a series of additional factors that are essential for telomere function. Although many shelterin-associated proteins have been so far identified, the inventory of shelterin-interacting factors required for telomere maintenance is still largely incomplete. Here, we characterize AKTIP/Ft1 (human AKTIP and mouse Ft1 are orthologous), a novel mammalian shelterin-bound factor identified on the basis of its homology with the Drosophila telomere protein Pendolino. AKTIP/Ft1 shares homology with the E2 variant ubiquitin-conjugating (UEV) enzymes and has been previously implicated in the control of apoptosis and in vesicle trafficking. RNAi-mediated depletion of AKTIP results in formation of telomere dysfunction foci (TIFs). Consistent with these results, AKTIP interacts with telomeric DNA and binds the shelterin components TRF1 and TRF2 both in vivo and in vitro. Analysis of AKTIP- depleted human primary fibroblasts showed that they are defective in PCNA recruiting and arrest in the S phase due to the activation of the intra S checkpoint. Accordingly, AKTIP physically interacts with PCNA and the RPA70 DNA replication factor. Ft1-depleted p53-/- MEFs did not arrest in the S phase but displayed significant increases in multiple telomeric signals (MTS) and sister telomere associations (STAs), two hallmarks of defective telomere replication. In addition, we found an epistatic relation for MST formation between Ft1 and TRF1, which has been previously shown to be required for replication fork progression through telomeric DNA. Ch-IP experiments further suggested that in AKTIP-depleted cells undergoing the S phase, TRF1 is less tightly bound to telomeric DNA than in controls. Thus, our results collectively suggest that AKTIP/Ft1 works in concert with TRF1 to facilitate telomeric DNA replication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Telomere-bound TRF1 and TRF2 stall the replication fork at telomeric repeats.Nucleic Acids Res. 2004 Mar 8;32(5):1627-37. doi: 10.1093/nar/gkh309. Print 2004. Nucleic Acids Res. 2004. PMID: 15007108 Free PMC article.
-
Post-translational modifications of TRF1 and TRF2 and their roles in telomere maintenance.Mech Ageing Dev. 2012 Jun;133(6):421-34. doi: 10.1016/j.mad.2012.05.002. Epub 2012 May 23. Mech Ageing Dev. 2012. PMID: 22634377 Review.
-
TRF1 negotiates TTAGGG repeat-associated replication problems by recruiting the BLM helicase and the TPP1/POT1 repressor of ATR signaling.Genes Dev. 2014 Nov 15;28(22):2477-91. doi: 10.1101/gad.251611.114. Epub 2014 Oct 24. Genes Dev. 2014. PMID: 25344324 Free PMC article.
-
Molecular recognition in complexes of TRF proteins with telomeric DNA.PLoS One. 2014 Feb 26;9(2):e89460. doi: 10.1371/journal.pone.0089460. eCollection 2014. PLoS One. 2014. PMID: 24586793 Free PMC article.
-
Shelterin: the protein complex that shapes and safeguards human telomeres.Genes Dev. 2005 Sep 15;19(18):2100-10. doi: 10.1101/gad.1346005. Genes Dev. 2005. PMID: 16166375 Review.
Cited by
-
Mammalian telomeres and their partnership with lamins.Nucleus. 2016 Apr 25;7(2):187-202. doi: 10.1080/19491034.2016.1179409. Nucleus. 2016. PMID: 27116558 Free PMC article. Review.
-
Drosophila Doublefault protein coordinates multiple events during male meiosis by controlling mRNA translation.Development. 2019 Nov 18;146(22):dev183053. doi: 10.1242/dev.183053. Development. 2019. PMID: 31645358 Free PMC article.
-
Genome-wide DNA methylation analysis in blood cells from patients with Werner syndrome.Clin Epigenetics. 2017 Aug 30;9:92. doi: 10.1186/s13148-017-0389-4. eCollection 2017. Clin Epigenetics. 2017. PMID: 28861129 Free PMC article.
-
p53-Sensitive Epileptic Behavior and Inflammation in Ft1 Hypomorphic Mice.Front Genet. 2018 Nov 27;9:581. doi: 10.3389/fgene.2018.00581. eCollection 2018. Front Genet. 2018. PMID: 30546381 Free PMC article.
-
Phase Separation in the Nucleus and at the Nuclear Periphery during Post-Mitotic Nuclear Envelope Reformation.Cells. 2022 May 25;11(11):1749. doi: 10.3390/cells11111749. Cells. 2022. PMID: 35681444 Free PMC article. Review.
References
-
- Blackburn E, Greider C, Szostak J. Telomeres and telomerase: the path from maize, Tetrahymena and yeast to human cancer and aging. Nat Med. 2006;12(10):1133–8. - PubMed
-
- Hockemeyer D, Palm W, Else T, Daniels J, Takai K, Ye J, et al. Telomere protection by mammalian Pot1 requires interaction with Tpp1. Nat Struct Mol Biol. 2007;14(8):754–61. - PubMed
-
- Denchi E, de Lange T. Protection of telomeres through independent control of ATM and ATR by TRF2 and POT1. Nature. 2007;448:1068–71. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
