

Modelling the mechanoreceptor's dynamic behaviour
- PMID: 26110655
- PMCID: PMC4523327
- DOI: 10.1111/joa.12328
Modelling the mechanoreceptor's dynamic behaviour
Abstract
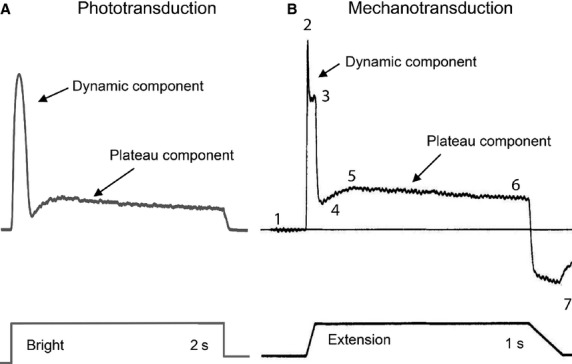
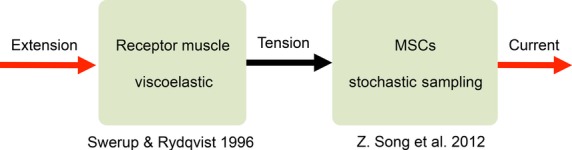
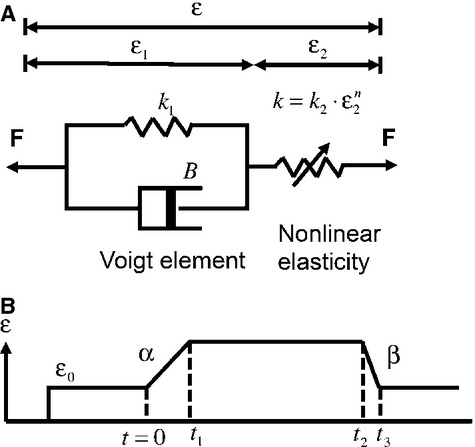
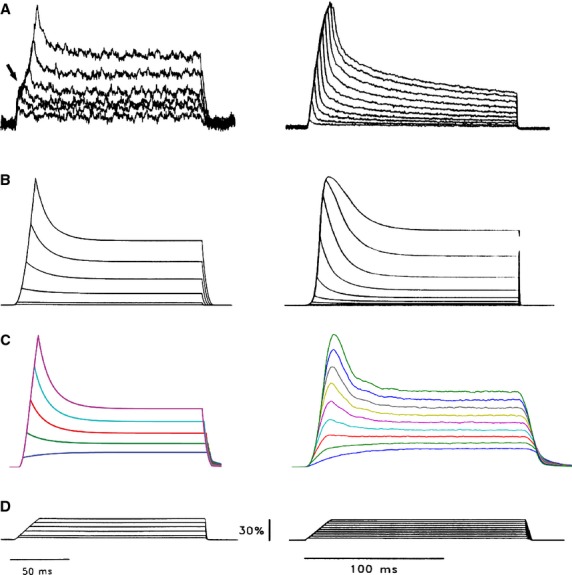
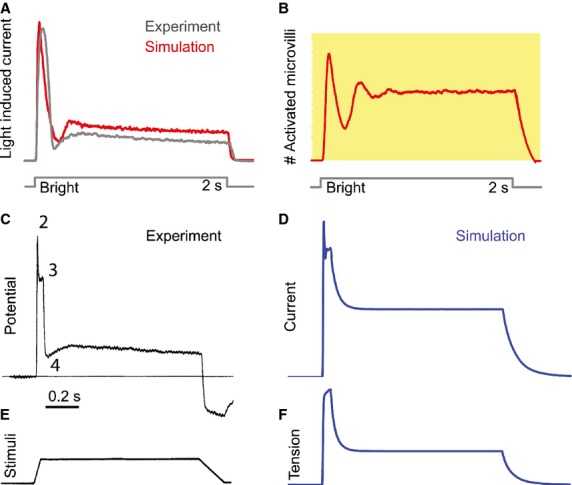
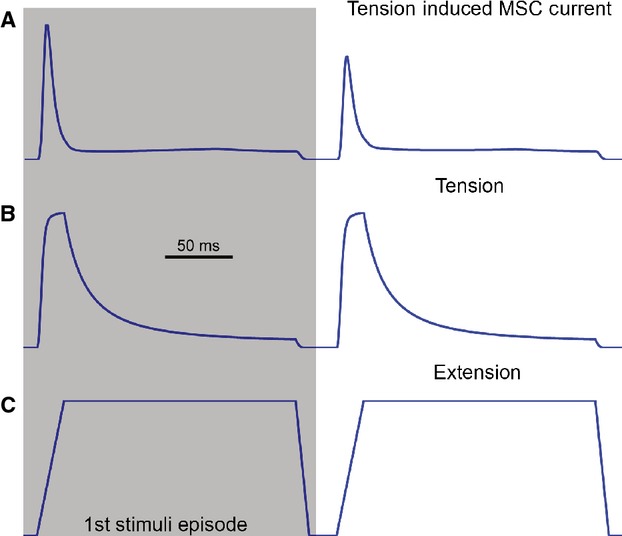
All sensory receptors adapt, i.e. they constantly adjust their sensitivity to external stimuli to match the current demands of the natural environment. Electrophysiological responses of sensory receptors from widely different modalities seem to exhibit common features related to adaptation, and these features can be used to examine the underlying sensory transduction mechanisms. Among the principal senses, mechanosensation remains the least understood at the cellular level. To gain greater insights into mechanosensory signalling, we investigated if mechanosensation displayed adaptive dynamics that could be explained by similar biophysical mechanisms in other sensory modalities. To do this, we adapted a fly photoreceptor model to describe the primary transduction process for a stretch-sensitive mechanoreceptor, taking into account the viscoelastic properties of the accessory muscle fibres and the biophysical properties of known mechanosensitive channels (MSCs). The model's output is in remarkable agreement with the electrical properties of a primary ending in an isolated decapsulated spindle; ramp-and-hold stretch evokes a characteristic pattern of potential change, consisting of a large dynamic depolarization during the ramp phase and a smaller static depolarization during the hold phase. The initial dynamic component is likely to be caused by a combination of the mechanical properties of the muscle fibres and a refractory state in the MSCs. Consistent with the literature, the current model predicts that the dynamic component is due to a rapid stress increase during the ramp. More novel predictions from the model are the mechanisms to explain the initial peak in the dynamic component. At the onset of the ramp, all MSCs are sensitive to external stimuli, but as they become refractory (inactivated), fewer MSCs are able to respond to the continuous stretch, causing a sharp decrease after the peak response. The same mechanism could contribute a faster component in the 'sensory habituation' of mechanoreceptors, in which a receptor responds more strongly to the first stimulus episode during repetitive stimulation.
Keywords: biophysical model; fly photoreceptor; refractory period; sensory adaptation; sensory habituation; stochastic adaptive sampling; stretch-sensitive mechanoreceptor.
© 2015 The Authors. Journal of Anatomy published by John Wiley & Sons Ltd on behalf of Anatomical Society.
Figures
Similar articles
-
A mathematical model of the crustacean stretch receptor neuron. Biomechanics of the receptor muscle, mechanosensitive ion channels, and macrotransducer properties.J Neurophysiol. 1996 Oct;76(4):2211-20. doi: 10.1152/jn.1996.76.4.2211. J Neurophysiol. 1996. PMID: 8899596
-
On the ionic mechanisms of adaptation in an isolated mechanoreceptor --an electrophysiological study.Acta Physiol Scand Suppl. 1983;520:1-43. Acta Physiol Scand Suppl. 1983. PMID: 6316733
-
[The fast and slow components of receptor adaptation in the discharge frequency of the primary muscle spindles in the cat].EEG EMG Z Elektroenzephalogr Elektromyogr Verwandte Geb. 1992 Mar;23(1):12-9. EEG EMG Z Elektroenzephalogr Elektromyogr Verwandte Geb. 1992. PMID: 1534049 German.
-
Stretching the imagination beyond muscle spindles - stretch-sensitive mechanisms in arthropods.J Anat. 2015 Aug;227(2):237-42. doi: 10.1111/joa.12329. Epub 2015 Jun 16. J Anat. 2015. PMID: 26076887 Free PMC article. Review.
-
Mechanotransduction in the muscle spindle.Pflugers Arch. 2015 Jan;467(1):175-90. doi: 10.1007/s00424-014-1536-9. Epub 2014 Jun 3. Pflugers Arch. 2015. PMID: 24888691 Free PMC article. Review.
Cited by
-
Assessment of the Effectiveness of Fascial Manipulation in Patients with Degenerative Disc Disease of the Lumbosacral Spine.Life (Basel). 2024 Dec 30;15(1):33. doi: 10.3390/life15010033. Life (Basel). 2024. PMID: 39859973 Free PMC article.
-
Viscoelastic Properties of Hyaluronan in Physiological Conditions.F1000Res. 2015 Aug 25;4:622. doi: 10.12688/f1000research.6885.1. eCollection 2015. F1000Res. 2015. PMID: 26594344 Free PMC article. Review.
-
Modeling elucidates how refractory period can provide profound nonlinear gain control to graded potential neurons.Physiol Rep. 2017 Jun;5(11):e13306. doi: 10.14814/phy2.13306. Physiol Rep. 2017. PMID: 28596301 Free PMC article.
-
Control of Mammalian Locomotion by Somatosensory Feedback.Compr Physiol. 2021 Dec 29;12(1):2877-2947. doi: 10.1002/cphy.c210020. Compr Physiol. 2021. PMID: 34964114 Free PMC article.
-
A biomimetic fly photoreceptor model elucidates how stochastic adaptive quantal sampling provides a large dynamic range.J Physiol. 2017 Aug 15;595(16):5439-5456. doi: 10.1113/JP273614. Epub 2017 May 17. J Physiol. 2017. PMID: 28369994 Free PMC article. Review.
References
-
- Boyd IA. The response of fast and slow nuclear bag fibres and nuclear chain fibres in isolated cat muscle spindles to fusimotor stimulation, and the effect of intrafusal contraction on the sensory endings. Q J Exp Physiol Cogn Med Sci. 1976;61:203–254. - PubMed
-
- Chalfie M. Neurosensory mechanotransduction. Nat Rev Mol Cell Biol. 2009;10:44–52. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
