

Structure-Function Analysis of the Non-Muscle Myosin Light Chain Kinase (nmMLCK) Isoform by NMR Spectroscopy and Molecular Modeling: Influence of MYLK Variants
- PMID: 26111161
- PMCID: PMC4482139
- DOI: 10.1371/journal.pone.0130515
Structure-Function Analysis of the Non-Muscle Myosin Light Chain Kinase (nmMLCK) Isoform by NMR Spectroscopy and Molecular Modeling: Influence of MYLK Variants
Abstract
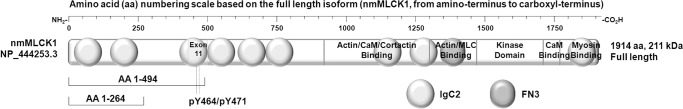
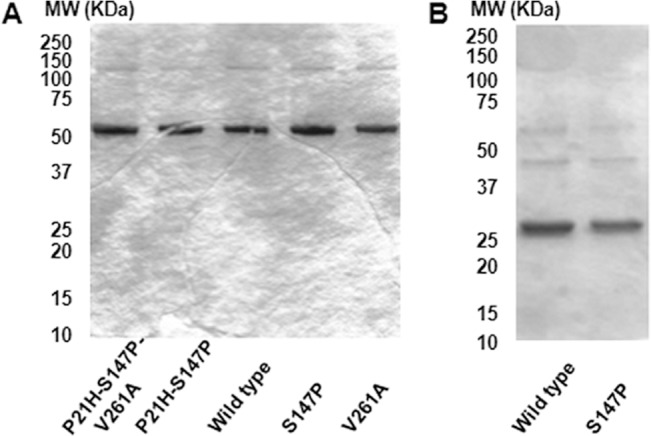
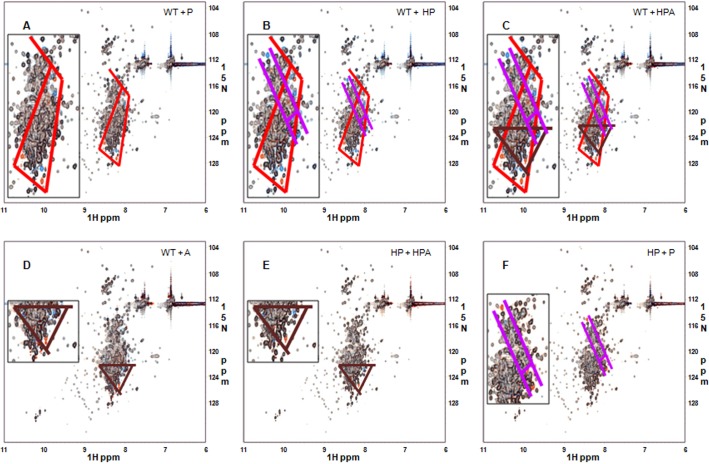
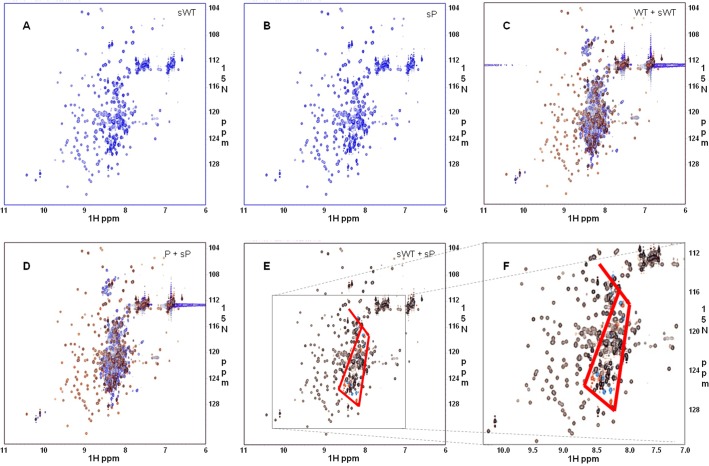
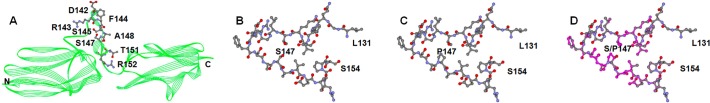
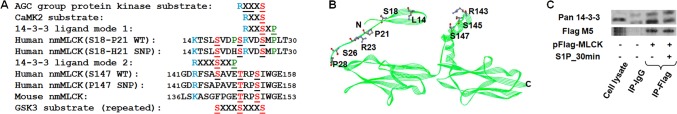
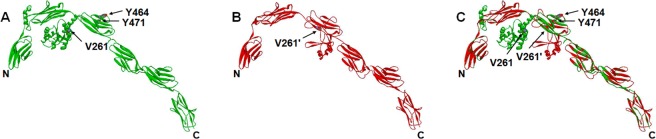
The MYLK gene encodes the multifunctional enzyme, myosin light chain kinase (MLCK), involved in isoform-specific non-muscle and smooth muscle contraction and regulation of vascular permeability during inflammation. Three MYLK SNPs (P21H, S147P, V261A) alter the N-terminal amino acid sequence of the non-muscle isoform of MLCK (nmMLCK) and are highly associated with susceptibility to acute lung injury (ALI) and asthma, especially in individuals of African descent. To understand the functional effects of SNP associations, we examined the N-terminal segments of nmMLCK by 1H-15N heteronuclear single quantum correlation (HSQC) spectroscopy, a 2-D NMR technique, and by in silico molecular modeling. Both NMR analysis and molecular modeling indicated SNP localization to loops that connect the immunoglobulin-like domains of nmMLCK, consistent with minimal structural changes evoked by these SNPs. Molecular modeling analysis identified protein-protein interaction motifs adversely affected by these MYLK SNPs including binding by the scaffold protein 14-3-3, results confirmed by immunoprecipitation and western blot studies. These structure-function studies suggest novel mechanisms for nmMLCK regulation, which may confirm MYLK as a candidate gene in inflammatory lung disease and advance knowledge of the genetic underpinning of lung-related health disparities.
Conflict of interest statement
Figures
Similar articles
-
Genetic and epigenetic regulation of the non-muscle myosin light chain kinase isoform by lung inflammatory factors and mechanical stress.Clin Sci (Lond). 2021 Apr 16;135(7):963-977. doi: 10.1042/CS20201448. Clin Sci (Lond). 2021. PMID: 33792658 Free PMC article.
-
An intronic MYLK variant associated with inflammatory lung disease regulates promoter activity of the smooth muscle myosin light chain kinase isoform.J Mol Med (Berl). 2012 Mar;90(3):299-308. doi: 10.1007/s00109-011-0820-9. Epub 2011 Oct 21. J Mol Med (Berl). 2012. PMID: 22015949
-
A MYLK variant regulates asthmatic inflammation via alterations in mRNA secondary structure.Eur J Hum Genet. 2015 Jun;23(6):874-6. doi: 10.1038/ejhg.2014.201. Epub 2014 Oct 1. Eur J Hum Genet. 2015. PMID: 25271083 Free PMC article.
-
Myosin light chain kinase as a multifunctional regulatory protein of smooth muscle contraction.IUBMB Life. 2001 Jun;51(6):337-44. doi: 10.1080/152165401753366087. IUBMB Life. 2001. PMID: 11758800 Review.
-
Phosphorylation of myosin regulatory light chain by myosin light chain kinase, and muscle contraction.Circ J. 2009 Feb;73(2):208-13. doi: 10.1253/circj.cj-08-1041. Epub 2008 Dec 26. Circ J. 2009. PMID: 19110504 Review.
Cited by
-
Myosin light chain kinase ( MYLK) coding polymorphisms modulate human lung endothelial cell barrier responses via altered tyrosine phosphorylation, spatial localization, and lamellipodial protrusions.Pulm Circ. 2018 Apr-Jun;8(2):2045894018764171. doi: 10.1177/2045894018764171. Epub 2018 Feb 26. Pulm Circ. 2018. PMID: 29480069 Free PMC article.
-
Mechanical Stress and Single Nucleotide Variants Regulate Alternative Splicing of the MYLK Gene.Am J Respir Cell Mol Biol. 2017 Jan;56(1):29-37. doi: 10.1165/rcmb.2016-0053OC. Am J Respir Cell Mol Biol. 2017. PMID: 27529643 Free PMC article.
-
Cardiomyocytes Sense Matrix Rigidity through a Combination of Muscle and Non-muscle Myosin Contractions.Dev Cell. 2018 Feb 5;44(3):326-336.e3. doi: 10.1016/j.devcel.2017.12.024. Epub 2018 Jan 26. Dev Cell. 2018. PMID: 29396114 Free PMC article.
-
A cortactin CTTN coding SNP contributes to lung vascular permeability and inflammatory disease severity in African descent subjects.Transl Res. 2022 Jun;244:56-74. doi: 10.1016/j.trsl.2022.02.002. Epub 2022 Feb 15. Transl Res. 2022. PMID: 35181549 Free PMC article.
-
Genomic and Genetic Approaches to Deciphering Acute Respiratory Distress Syndrome Risk and Mortality.Antioxid Redox Signal. 2019 Nov 10;31(14):1027-1052. doi: 10.1089/ars.2018.7701. Epub 2019 Jun 18. Antioxid Redox Signal. 2019. PMID: 31016989 Free PMC article. Review.
References
-
- Dudek SM, Garcia JGN. Cytoskeletal regulation of pulmonary vascular permeability. J Appl Physiol. 2001;91(4):1487–500. - PubMed
-
- Kamm KE, Stull JT. Dedicated myosin light chain kinases with diverse cellular functions. J Biol Chem. 2001;276(7):4527–30. - PubMed
-
- Garcia JGN, Verin AD, Herenyiova M, English D. Adherent neutrophils activate endothelial myosin light chain kinase: role in transendothelial migration. J Appl Physiol. 1998;84(5):1817–21. - PubMed
-
- Saito H, Minamiya Y, Kitamura M, Saito S, Enomoto K, Terada K, et al. Endothelial myosin light chain kinase regulates neutrophil migration across human umbilical vein endothelial cell monolayer. J Immunol. 1998;161(3):1533–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
