

Endothelial MMP14 is required for endothelial-dependent growth support of human airway basal cells
- PMID: 26116571
- PMCID: PMC4541042
- DOI: 10.1242/jcs.168179
Endothelial MMP14 is required for endothelial-dependent growth support of human airway basal cells
Abstract
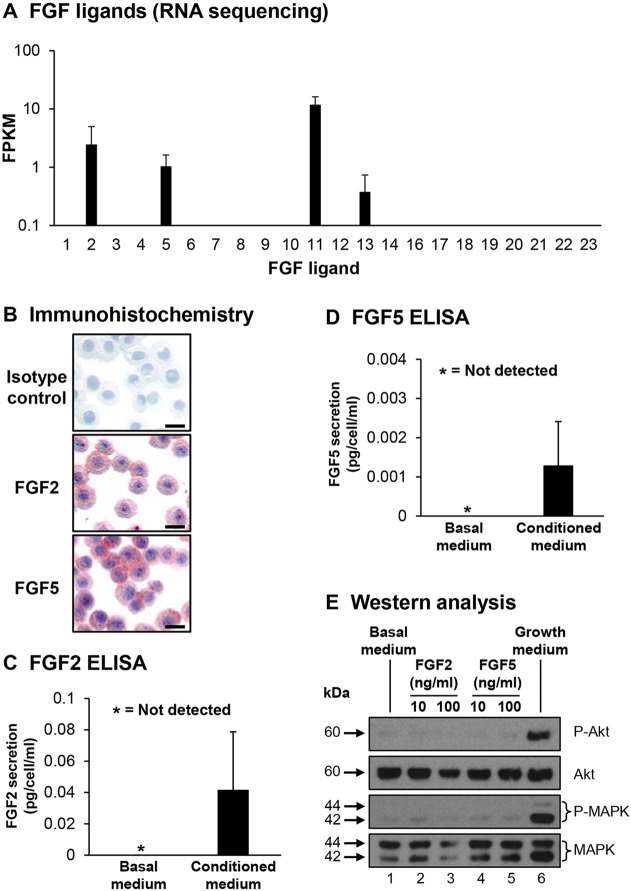
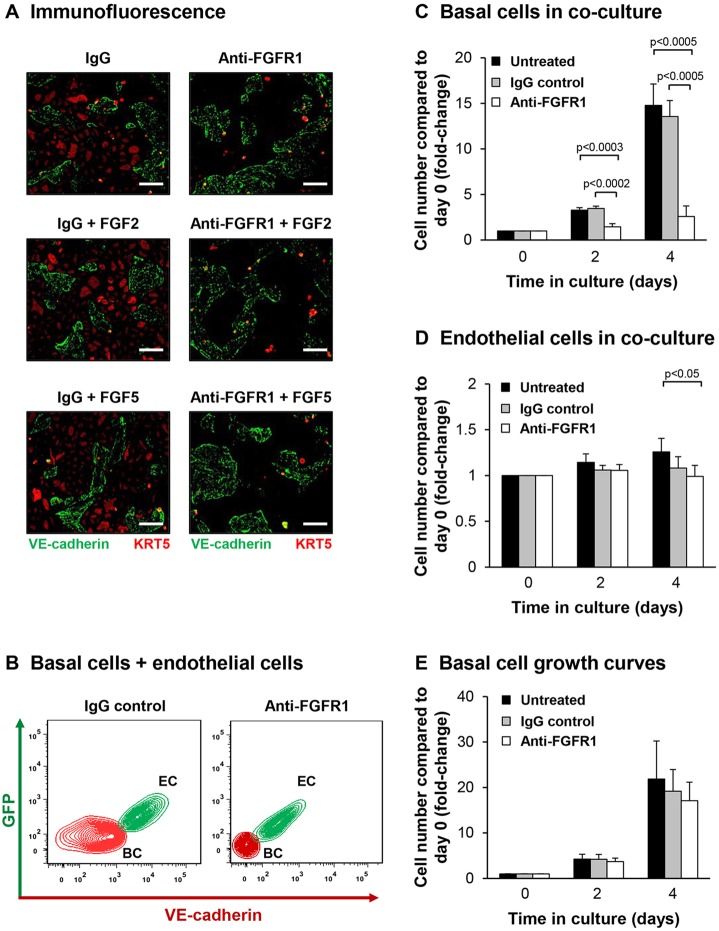
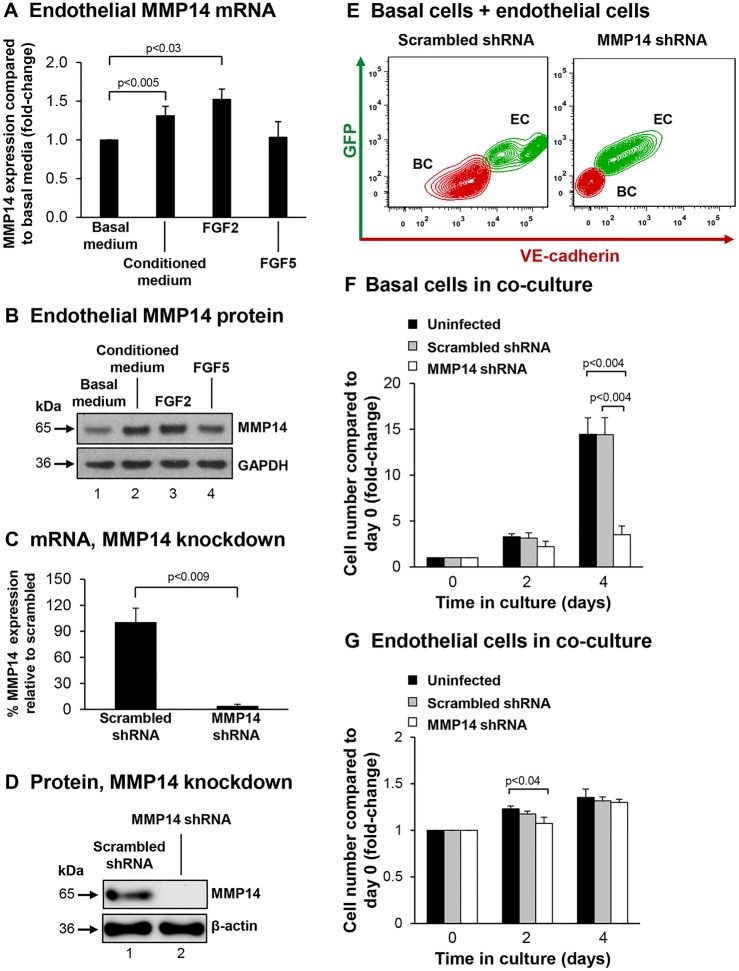
Human airway basal cells are the stem (or progenitor) population of the airway epithelium, and play a central role in anchoring the epithelium to the basement membrane. The anatomic position of basal cells allows for potential paracrine signaling between them and the underlying non-epithelial stromal cells. In support of this, we have previously demonstrated that endothelial cells support growth of basal cells during co-culture through vascular endothelial growth factor A (VEGFA)-mediated signaling. Building on these findings, we found, by RNA sequencing analysis, that basal cells expressed multiple fibroblast growth factor (FGF) ligands (FGF2, FGF5, FGF11 and FGF13) and that only FGF2 and FGF5 were capable of functioning in a paracrine manner to activate classical FGF receptor (FGFR) signaling. Antibody-mediated blocking of FGFR1 during basal-cell-endothelial-cell co-culture significantly reduced the endothelial-cell-dependent basal cell growth. Stimulation of endothelial cells with basal-cell-derived growth factors induced endothelial cell expression of matrix metallopeptidase 14 (MMP14), and short hairpin RNA (shRNA)-mediated knockdown of endothelial cell MMP14 significantly reduced the endothelial-cell-dependent growth of basal cells. Overall, these data characterize a new growth-factor-mediated reciprocal 'crosstalk' between human airway basal cells and endothelial cells that regulates proliferation of basal cells.
Keywords: Airway basal cell; Crosstalk; Endothelial cell; MMP14; Progenitor cell.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare that they have no competing interests, but notify that S.R. is the founder of and consultant to Angiocrine Bioscience New York, NY, USA.
Figures
References
-
- Allerstorfer S., Sonvilla G., Fischer H., Spiegl-Kreinecker S., Gauglhofer C., Setinek U., Czech T., Marosi C., Buchroithner J., Pichler J. et al. (2008). FGF5 as an oncogenic factor in human glioblastoma multiforme: autocrine and paracrine activities. Oncogene 27, 4180-4190. 10.1038/onc.2008.61 - DOI - PMC - PubMed
-
- Curradi G., Walters M. S., Ding B.-S., Rafii S., Hackett N. R. and Crystal R. G. (2012). Airway basal cell vascular endothelial growth factor-mediated cross-talk regulates endothelial cell-dependent growth support of human airway basal cells. Cell. Mol. Life Sci. 69, 2217-2231. 10.1007/s00018-012-0922-8 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
