

Specifying and protecting germ cell fate
- PMID: 26122616
- PMCID: PMC4698964
- DOI: 10.1038/nrm4009
Specifying and protecting germ cell fate
Abstract
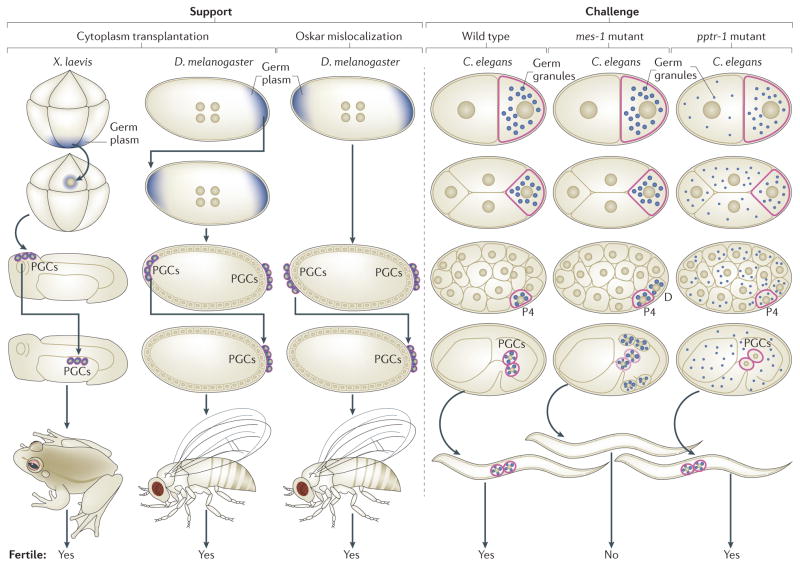
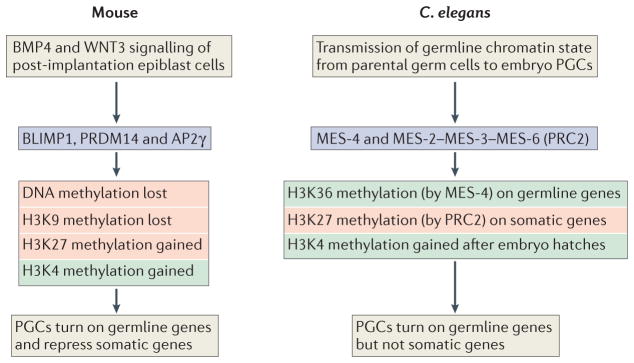
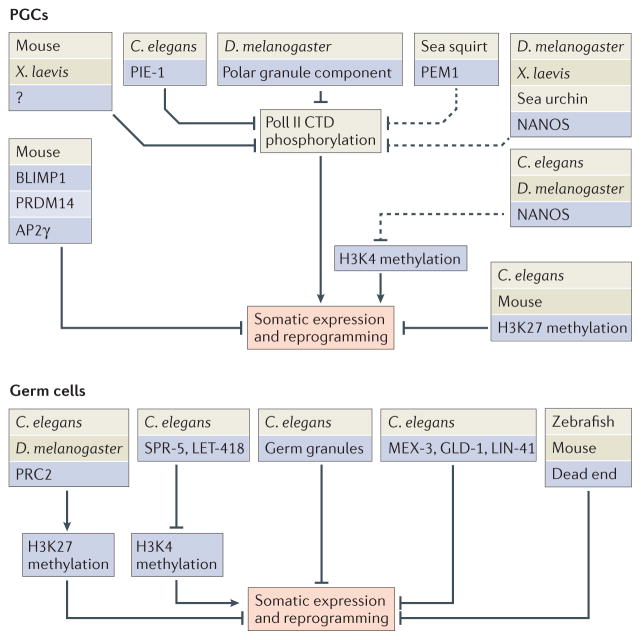
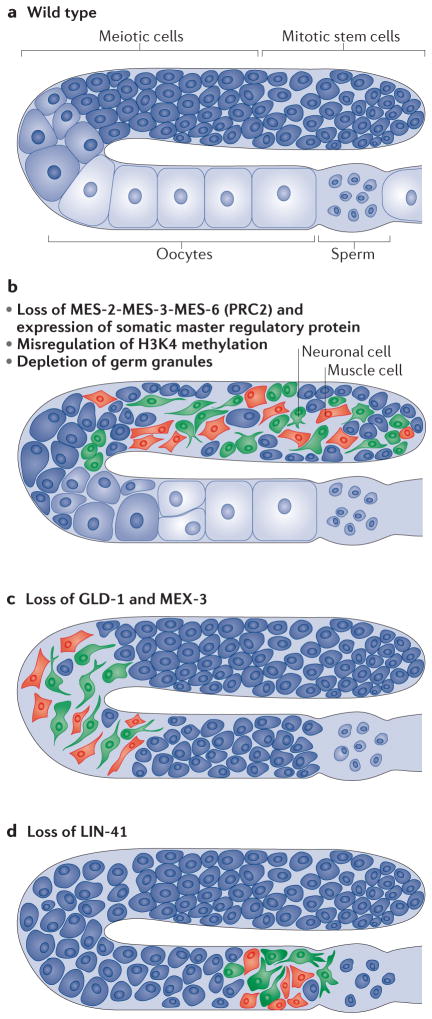
Germ cells are the special cells in the body that undergo meiosis to generate gametes and subsequently entire new organisms after fertilization, a process that continues generation after generation. Recent studies have expanded our understanding of the factors and mechanisms that specify germ cell fate, including the partitioning of maternally supplied 'germ plasm', inheritance of epigenetic memory and expression of transcription factors crucial for primordial germ cell (PGC) development. Even after PGCs are specified, germline fate is labile and thus requires protective mechanisms, such as global transcriptional repression, chromatin state alteration and translation of only germline-appropriate transcripts. Findings from diverse species continue to provide insights into the shared and divergent needs of these special reproductive cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Seydoux G, Braun RE. Pathway to totipotency: lessons from germ cells. Cell. 2006;127:891–904. - PubMed
-
- Hegner RW. Effects of removing the germ-cell determinants from the eggs of some chrysomelid beetles. Biol Bull. 1908;16:19–26.
-
- Eddy EM. Germ plasm and the differentiation of the germ cell line. Int Rev Cytol. 1975;43:229–280. - PubMed
-
- Strome S, Wood WB. Generation of asymmetry and segregation of germ-line granules in early C elegans embryos. Cell. 1983;35:15–25. - PubMed
-
- Hay B, Jan LY, Jan YN. A protein component of Drosophila polar granules is encoded by vasa and has extensive sequence similarity to ATP-dependent helicases. Cell. 1988;55:577–587. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
