

Enzymatic measurement of phosphatidylglycerol and cardiolipin in cultured cells and mitochondria
- PMID: 26122953
- PMCID: PMC4485230
- DOI: 10.1038/srep11737
Enzymatic measurement of phosphatidylglycerol and cardiolipin in cultured cells and mitochondria
Abstract
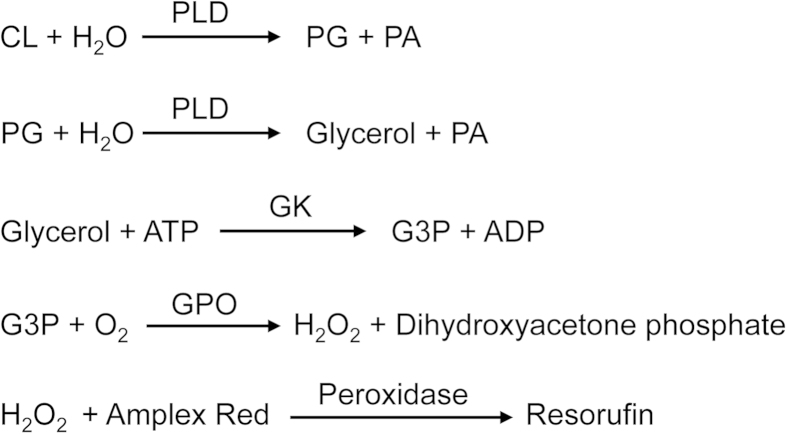
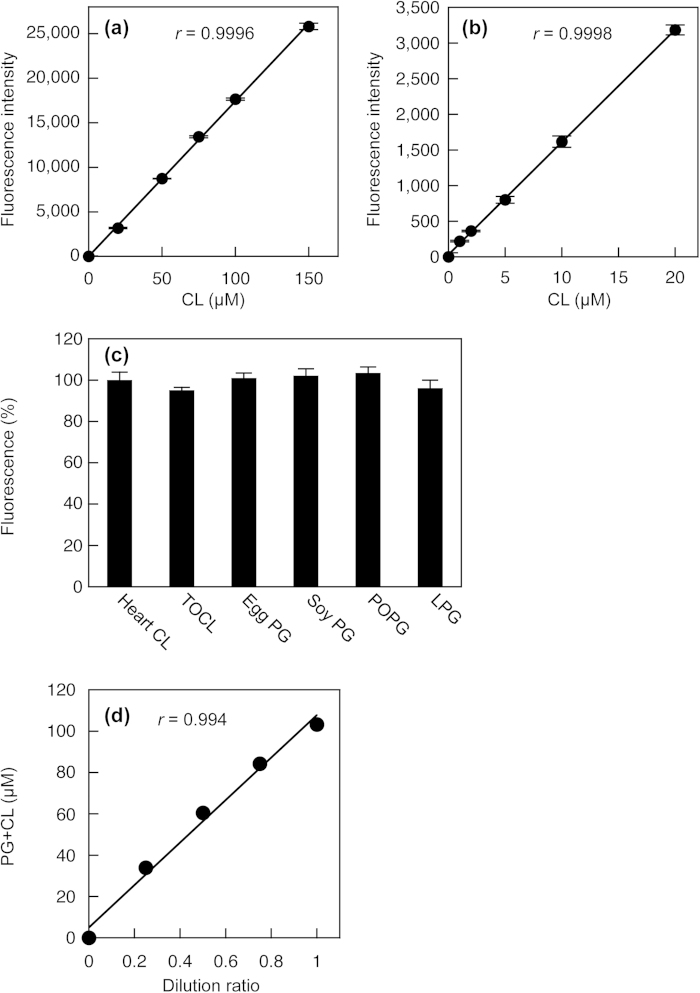
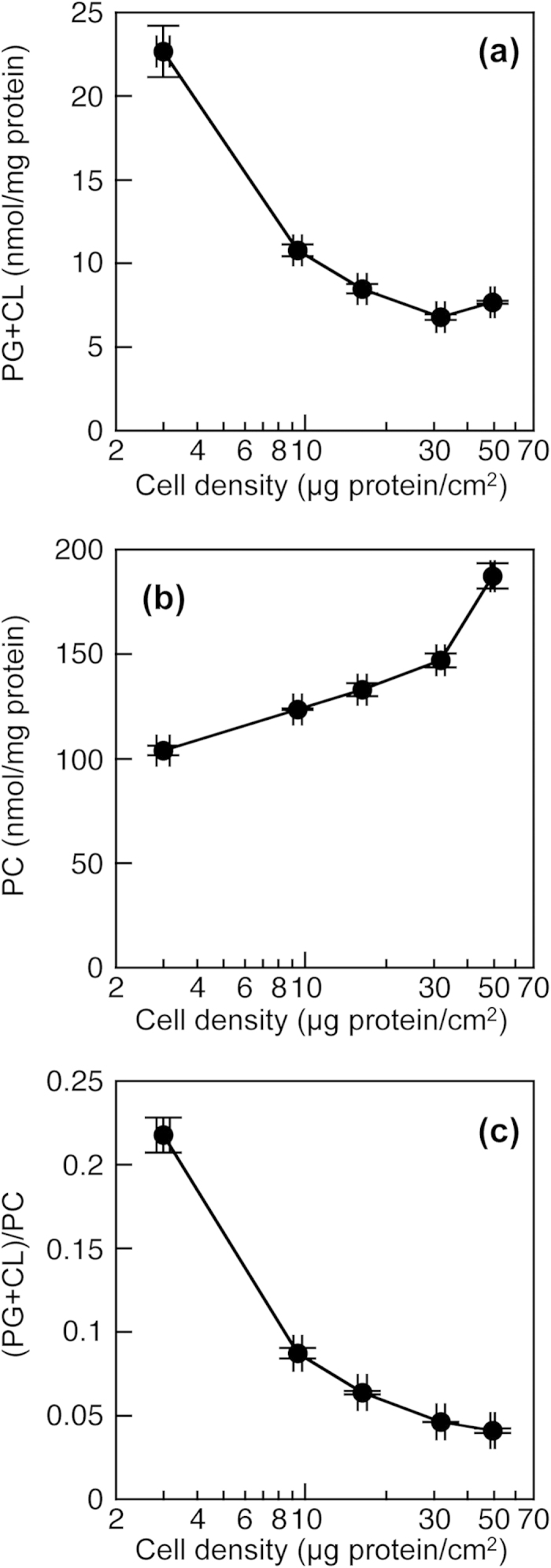
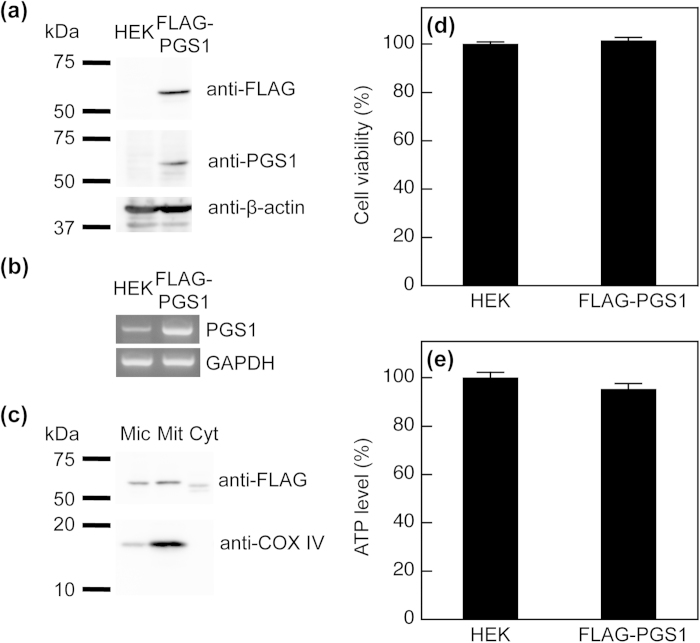
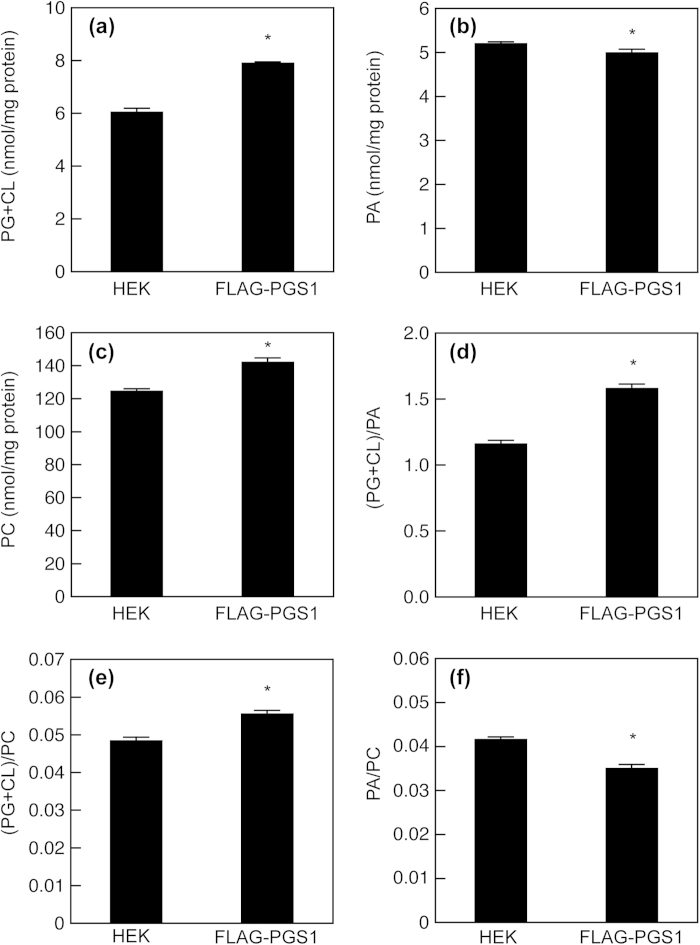
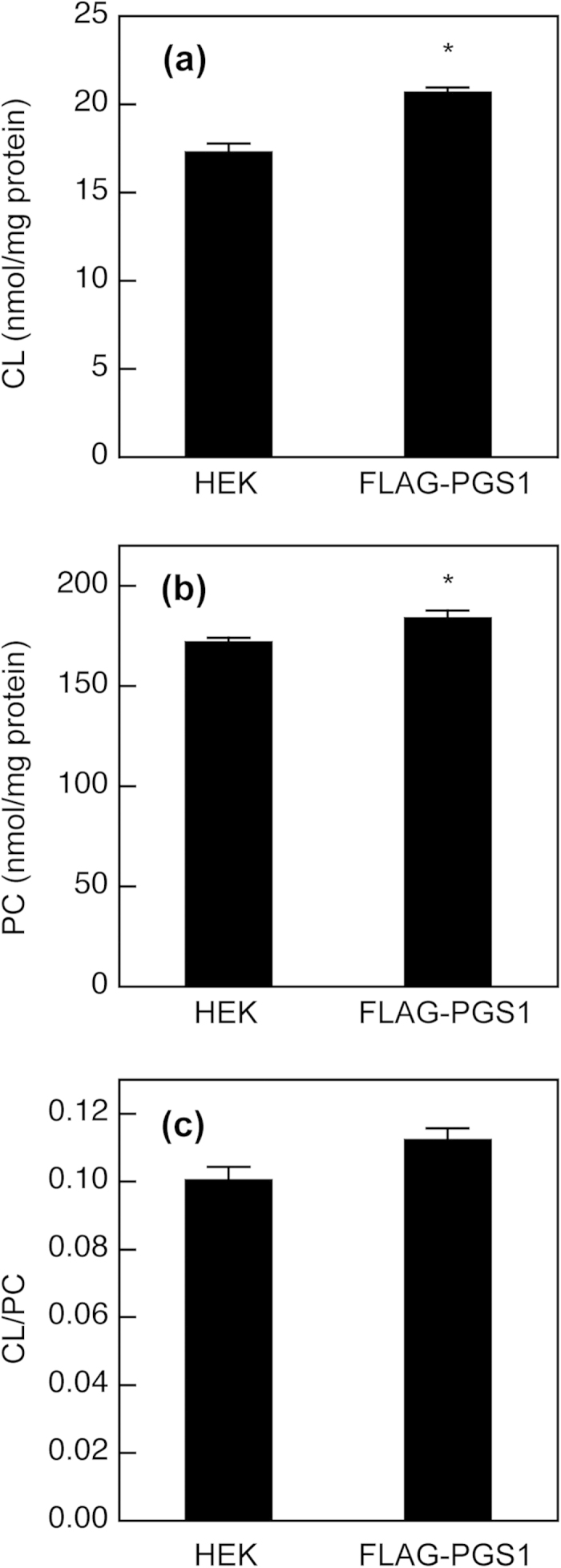
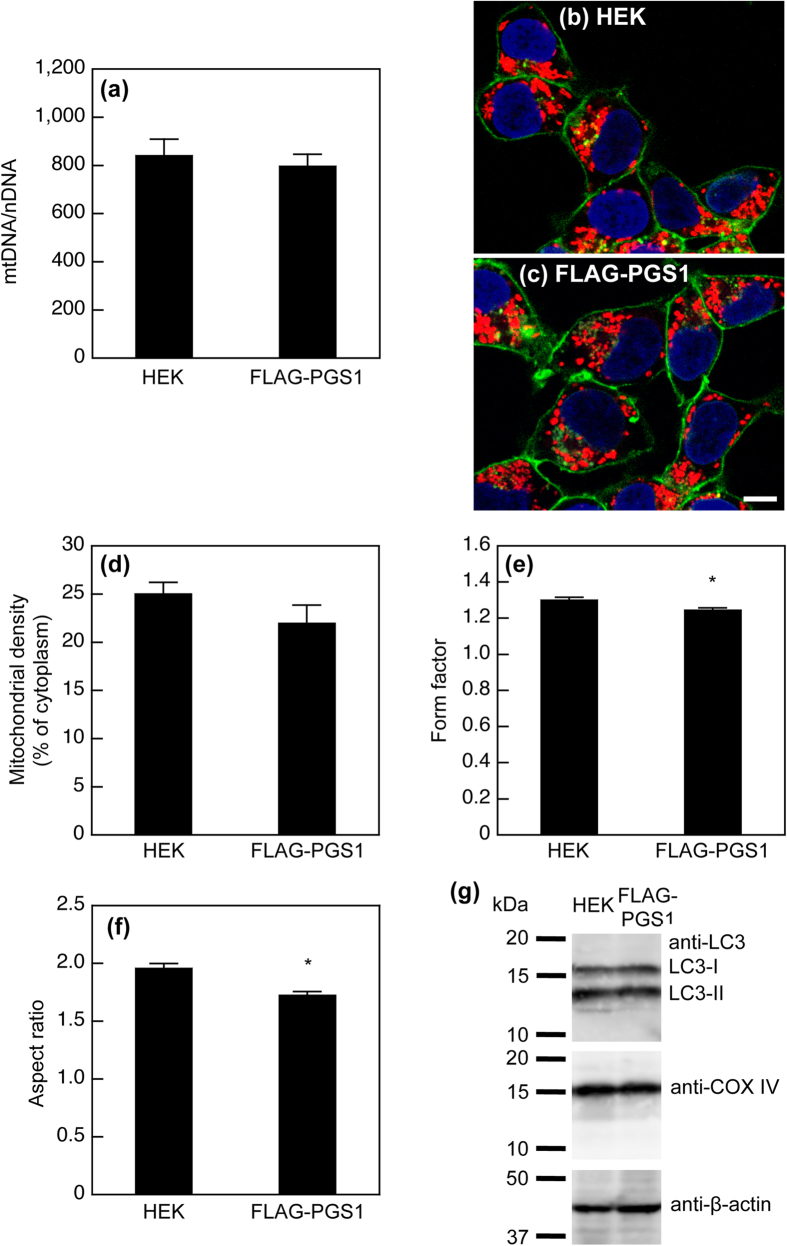
Phosphatidylglycerol (PG) and cardiolipin (CL) are synthesized in mitochondria and regulate numerous biological functions. In this study, a novel fluorometric method was developed for measuring PG and CL using combinations of specific enzymes and Amplex Red. This assay quantified the sum of PG and CL (PG + CL) regardless of the species of fatty acyl chain. The calibration curve for PG + CL measurement was linear, and the detection limit was 1 μM (10 pmol in the reaction mixture). This new method was applied to the determinations of PG + CL content in HEK293 cells and CL content in purified mitochondria, because the mitochondrial content of PG is negligible compared with that of CL. We demonstrated that the PG+CL content was greater at low cell density than at high cell density. The overexpression of phosphatidylglycerophosphate synthase 1 (PGS1) increased the cellular contents of PG + CL and phosphatidylcholine (PC), and reduced that of phosphatidic acid. PGS1 overexpression also elevated the mitochondrial contents of CL and PC, but had no effect on the number of mitochondria per cell. In addition to the enzymatic measurements of other phospholipids, this simple, sensitive and high-throughput assay for measuring PG + CL can be used to understand cellular, physiological and pathological processes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Daum G. & Vance J. E. Import of lipids into mitochondria. Prog. Lipid Res. 36, 103–130 (1997). - PubMed
-
- Horvath S. E. & Daum G. Lipids of mitochondria. Prog. Lipid Res. 52, 590–614 (2013). - PubMed
-
- Touchstone J. C., Alvarez J. G., Levin S. S. & Storey B. T. Evidence for diplasmalogen as the major component of rabbit sperm phosphatidylethanolamine. Lipids 20, 869–875 (1985). - PubMed
-
- Wada H. & Murata N. The essential role of phosphatidylglycerol in photosynthesis. Photosynth Res 92, 205–215 (2007). - PubMed
-
- de Kroon A. I., Dolis D., Mayer A., Lill R. & de Kruijff B. Phospholipid composition of highly purified mitochondrial outer membranes of rat liver and Neurospora crassa. Is cardiolipin present in the mitochondrial outer membrane? Biochim. Biophys. Acta 1325, 108–116 (1997). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
