

Identification of cis-suppression of human disease mutations by comparative genomics
- PMID: 26123021
- PMCID: PMC4537371
- DOI: 10.1038/nature14497
Identification of cis-suppression of human disease mutations by comparative genomics
Abstract
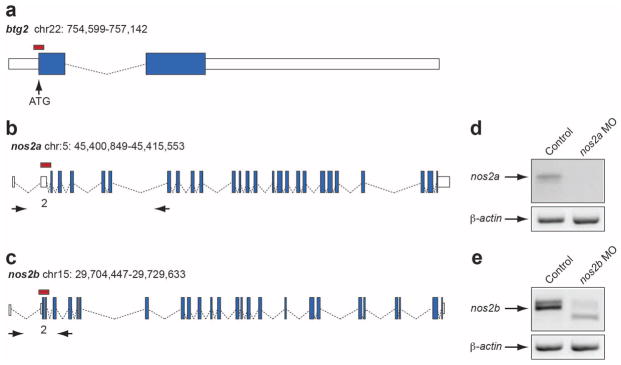
Patterns of amino acid conservation have served as a tool for understanding protein evolution. The same principles have also found broad application in human genomics, driven by the need to interpret the pathogenic potential of variants in patients. Here we performed a systematic comparative genomics analysis of human disease-causing missense variants. We found that an appreciable fraction of disease-causing alleles are fixed in the genomes of other species, suggesting a role for genomic context. We developed a model of genetic interactions that predicts most of these to be simple pairwise compensations. Functional testing of this model on two known human disease genes revealed discrete cis amino acid residues that, although benign on their own, could rescue the human mutations in vivo. This approach was also applied to ab initio gene discovery to support the identification of a de novo disease driver in BTG2 that is subject to protective cis-modification in more than 50 species. Finally, on the basis of our data and models, we developed a computational tool to predict candidate residues subject to compensation. Taken together, our data highlight the importance of cis-genomic context as a contributor to protein evolution; they provide an insight into the complexity of allele effect on phenotype; and they are likely to assist methods for predicting allele pathogenicity.
Conflict of interest statement
The authors declare no competing financial interests. Readers are welcome to comment on the online version of the paper.
Figures







References
-
- Cooper GM, Shendure J. Needles in stacks of needles: finding disease-causal variants in a wealth of genomic data. Nature Rev Genet. 2011;12:628–640. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM078598/GM/NIGMS NIH HHS/United States
- R01 EY021872/EY/NEI NIH HHS/United States
- R01 DK072301/DK/NIDDK NIH HHS/United States
- R01 DK095721/DK/NIDDK NIH HHS/United States
- P50 DK096415/DK/NIDDK NIH HHS/United States
- R00 HG007229/HG/NHGRI NIH HHS/United States
- U01 HG006500/HG/NHGRI NIH HHS/United States
- R01 DK075972/DK/NIDDK NIH HHS/United States
- R01EY021872/EY/NEI NIH HHS/United States
- R01 HD042601/HD/NICHD NIH HHS/United States
- R01 MH101244/MH/NIMH NIH HHS/United States
- R01DK075972/DK/NIDDK NIH HHS/United States
- R01HD04260/HD/NICHD NIH HHS/United States
- R01DK072301/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
