

A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana
- PMID: 26124115
- PMCID: PMC4507220
- DOI: 10.1073/pnas.1508432112
A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana
Abstract
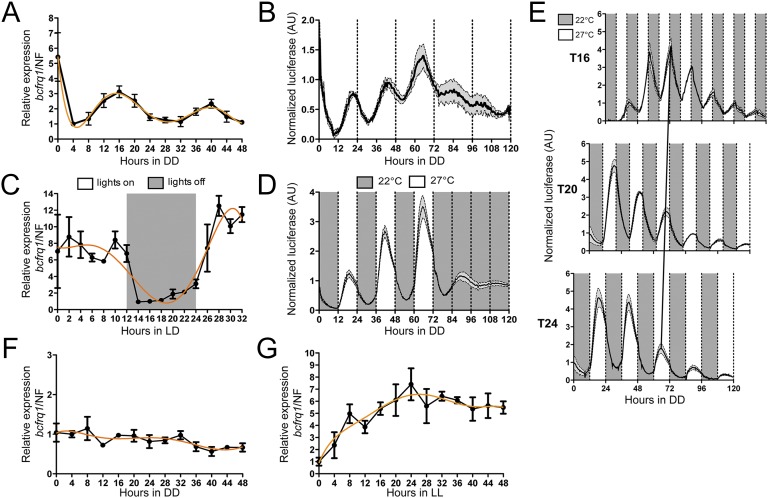
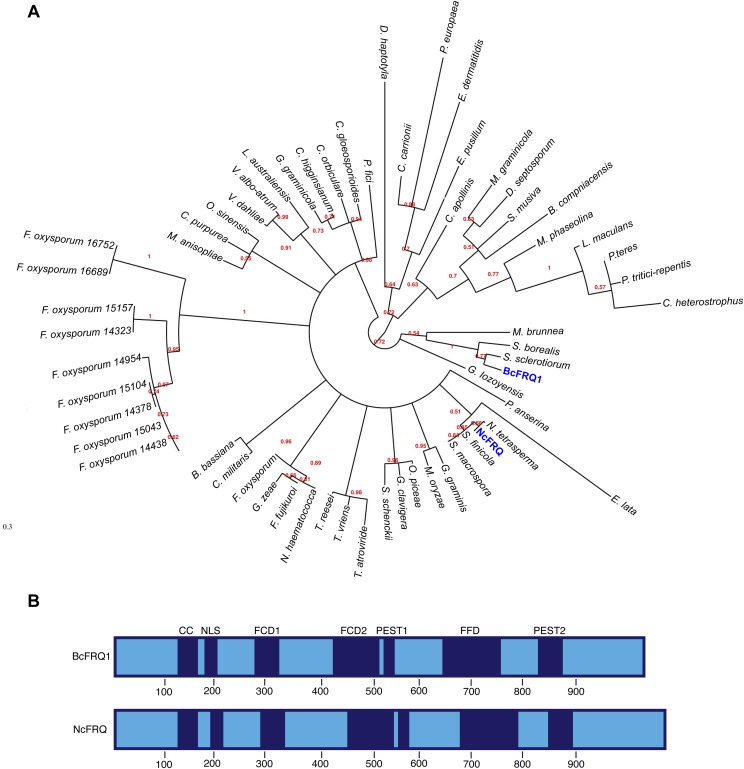
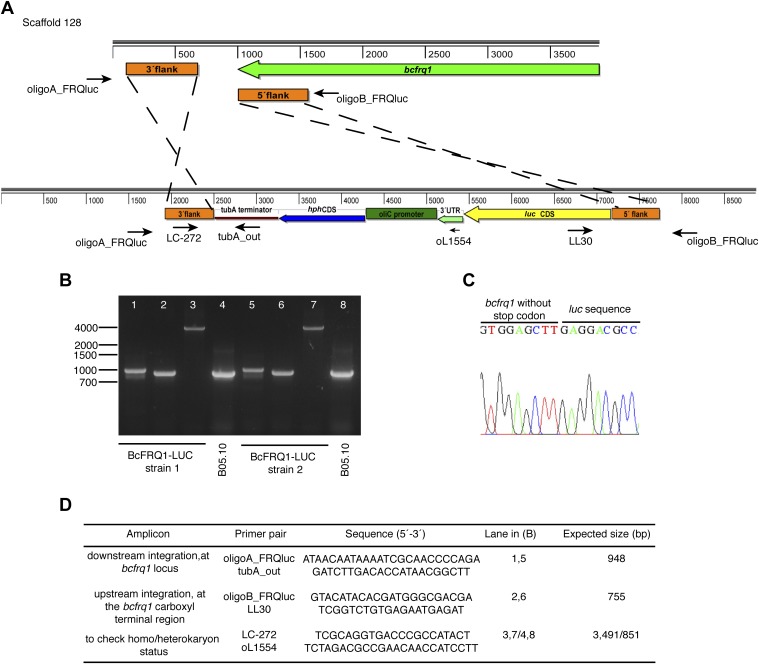
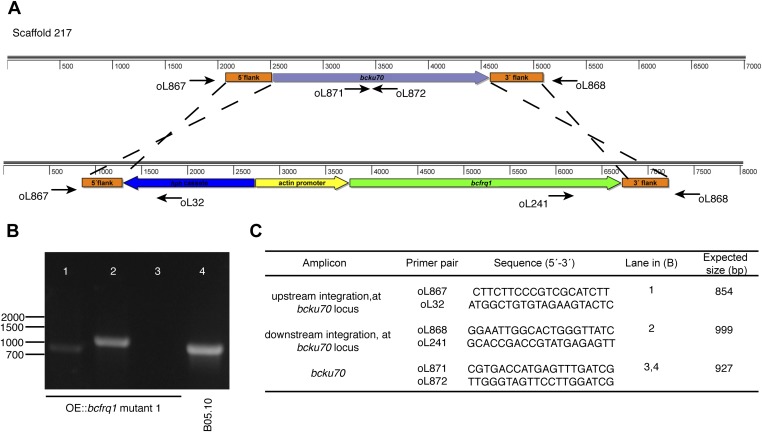
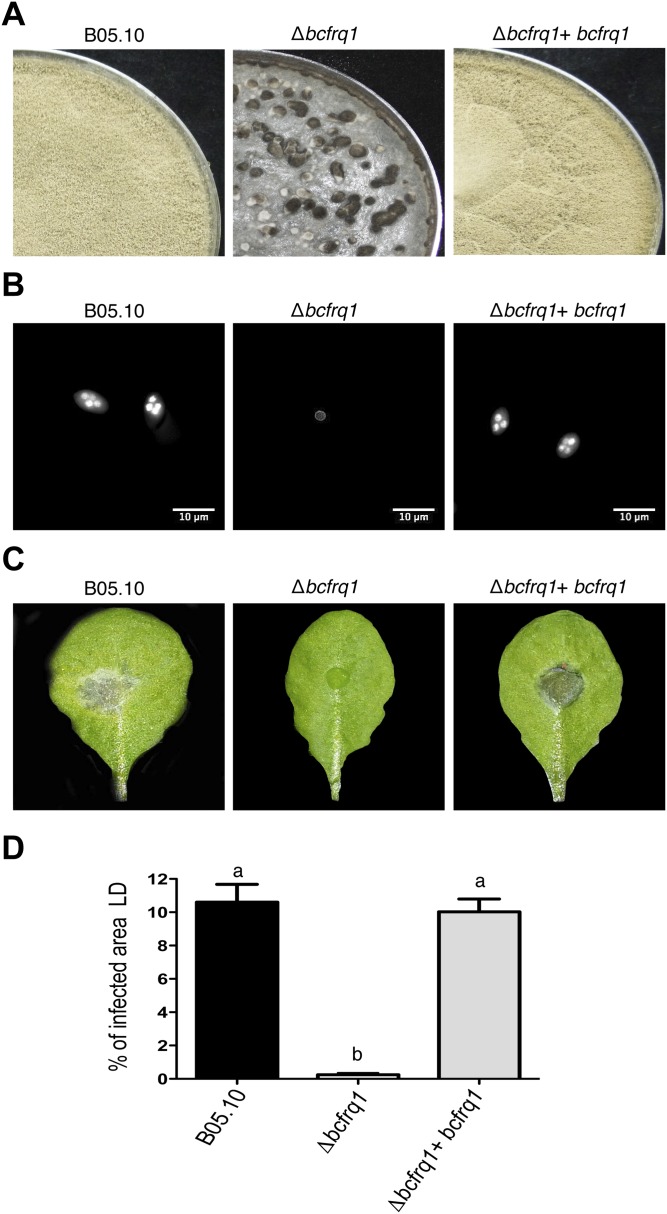
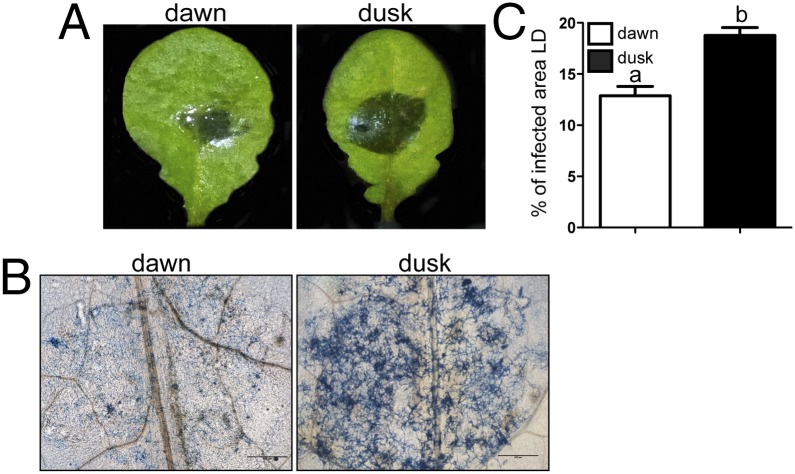
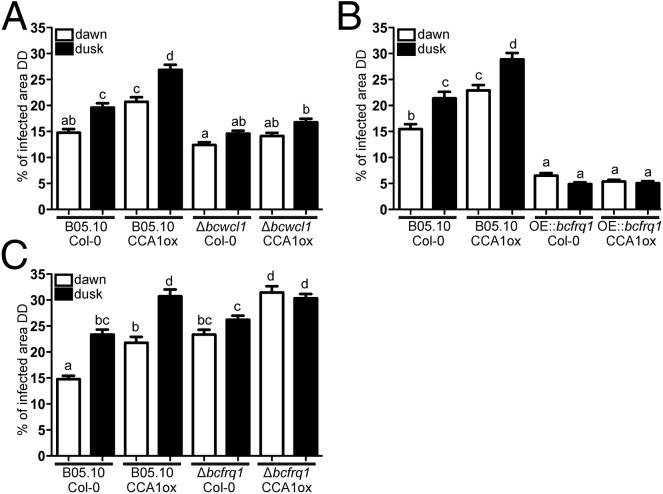
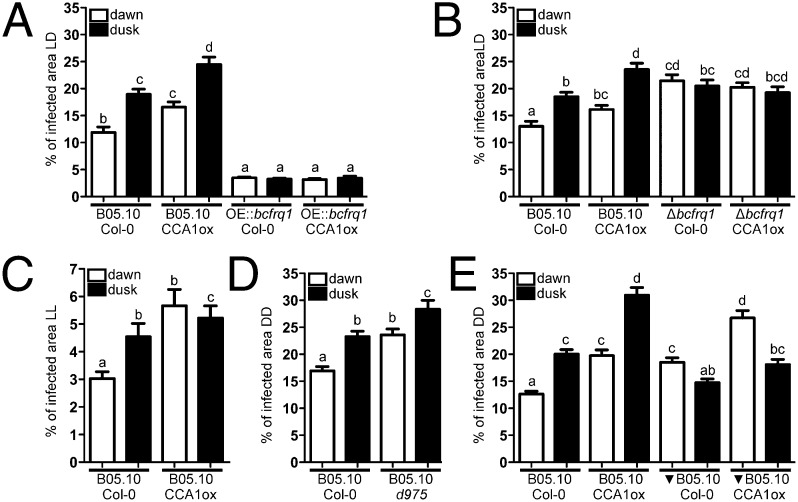
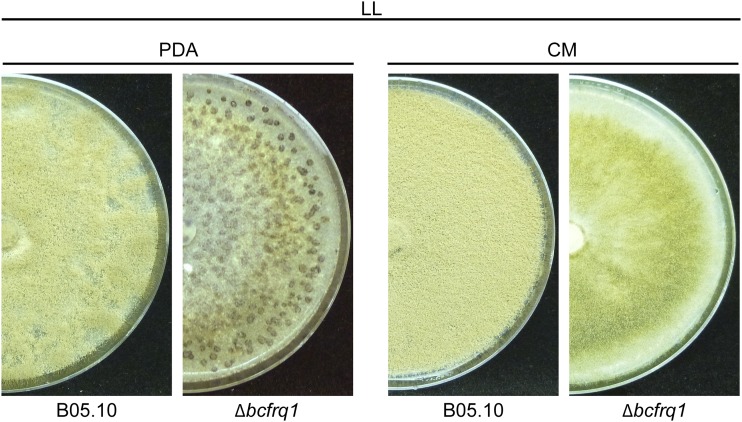
The circadian clock of the plant model Arabidopsis thaliana modulates defense mechanisms impacting plant-pathogen interactions. Nevertheless, the effect of clock regulation on pathogenic traits has not been explored in detail. Moreover, molecular description of clocks in pathogenic fungi--or fungi in general other than the model ascomycete Neurospora crassa--has been neglected, leaving this type of question largely unaddressed. We sought to characterize, therefore, the circadian system of the plant pathogen Botrytis cinerea to assess if such oscillatory machinery can modulate its virulence potential. Herein, we show the existence of a functional clock in B. cinerea, which shares similar components and circuitry with the Neurospora circadian system, although we found that its core negative clock element FREQUENCY (BcFRQ1) serves additional roles, suggesting extracircadian functions for this protein. We observe that the lesions produced by this necrotrophic fungus on Arabidopsis leaves are smaller when the interaction between these two organisms occurs at dawn. Remarkably, this effect does not depend solely on the plant clock, but instead largely relies on the pathogen circadian system. Genetic disruption of the B. cinerea oscillator by mutation, overexpression of BcFRQ1, or by suppression of its rhythmicity by constant light, abrogates circadian regulation of fungal virulence. By conducting experiments with out-of-phase light:dark cycles, we confirm that indeed, it is the fungal clock that plays the main role in defining the outcome of the Arabidopsis-Botrytis interaction, providing to our knowledge the first evidence of a microbial clock modulating pathogenic traits at specific times of the day.
Keywords: Arabidopsis thaliana; Botrytis cinerea; circadian clock; plant–pathogen interaction; virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
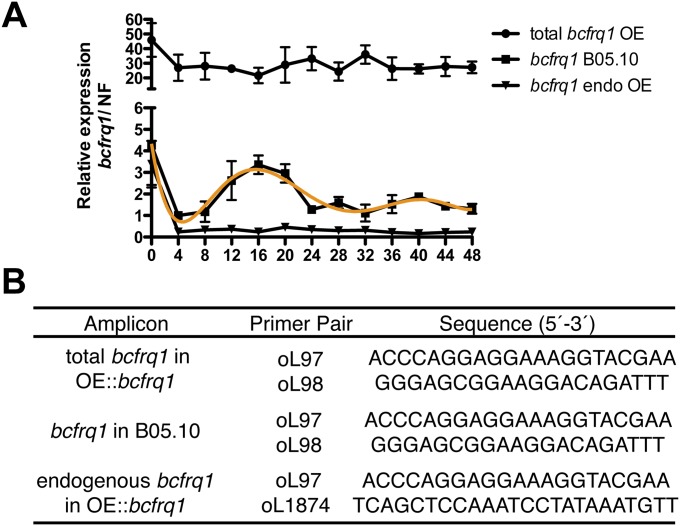
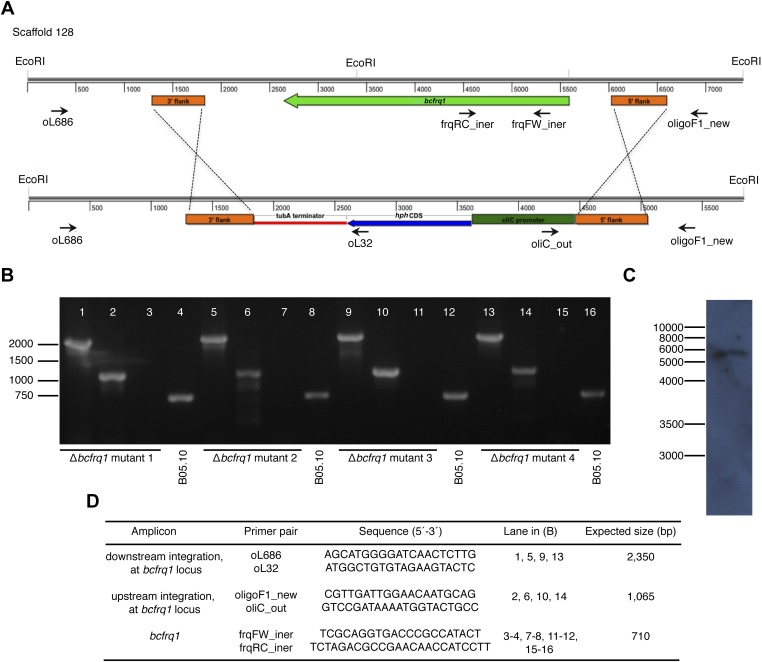
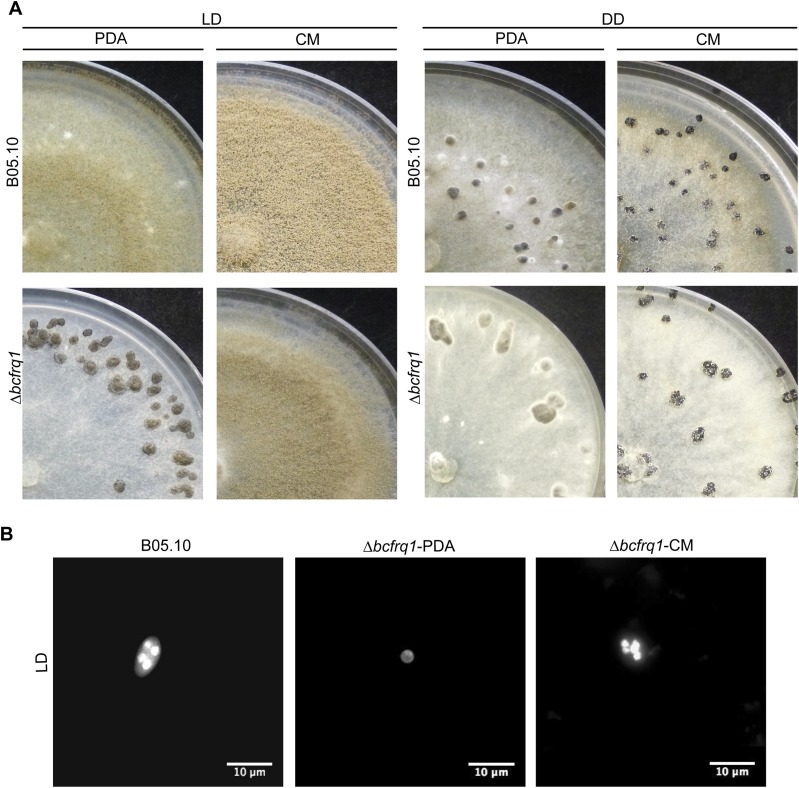
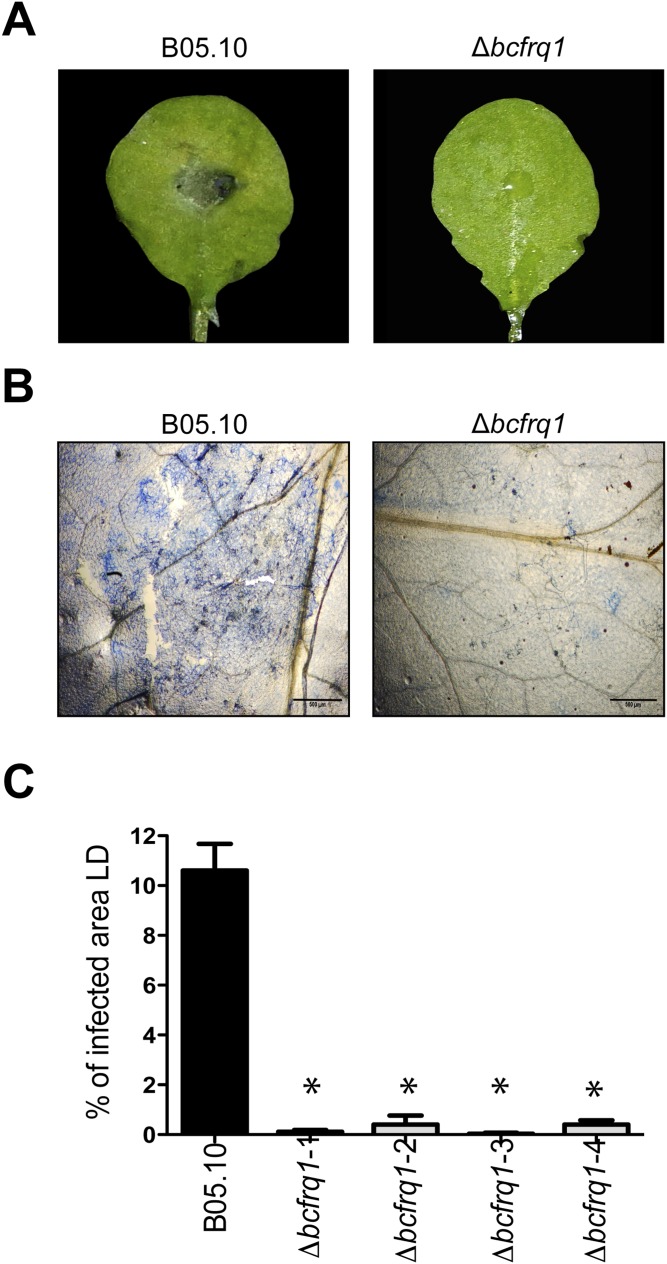

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
