

Resistance of R-Ras knockout mice to skin tumour induction
- PMID: 26133397
- PMCID: PMC4488886
- DOI: 10.1038/srep11663
Resistance of R-Ras knockout mice to skin tumour induction
Abstract
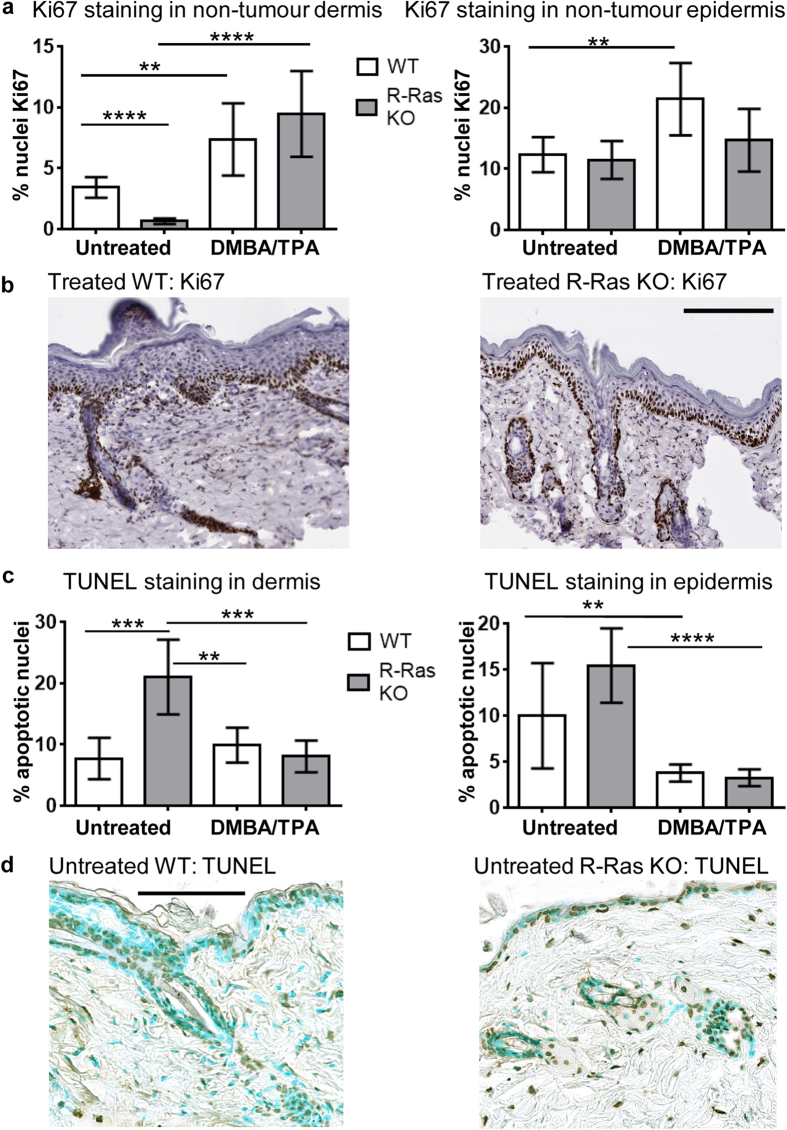
The R-ras gene encodes a small GTPase that is a member of the Ras family. Despite close sequence similarities, R-Ras is functionally distinct from the prototypic Ras proteins; no transformative activity and no activating mutations of R-Ras in human malignancies have been reported for it. R-Ras activity appears inhibitory towards tumour proliferation and invasion, and to promote cellular quiescence. Contrary to this, using mice with a deletion of the R-ras gene, we found that R-Ras facilitates DMBA/TPA-induced skin tumour induction. The tumours appeared in wild-type (WT) mice on average 6 weeks earlier than in R-Ras knockout (R-Ras KO) mice. WT mice developed almost 6 times more tumours than R-Ras KO mice. Despite strong R-Ras protein expression in the dermal blood vessels, no R-Ras could be detected in the epidermis from where the tumours arose. The DMBA/TPA skin tumourigenesis-model is highly dependent upon inflammation, and we found a greatly attenuated skin inflammatory response to DMBA/TPA-treatment in the R-Ras KO mice in the context of leukocyte infiltration and proinflammatory cytokine expression. Thus, these data suggest that despite its characterised role in promoting cellular quiescence, R-Ras is pro-tumourigenic in the DMBA/TPA tumour model and important for the inflammatory response to DMBA/TPA treatment.
Figures




Similar articles
-
Genetic alterations cooperate with v-Ha-ras to accelerate multistage carcinogenesis in TG.AC transgenic mouse skin.Cancer Res. 1995 Jul 15;55(14):3171-8. Cancer Res. 1995. PMID: 7606738
-
Only a subset of 12-O-tetradecanoylphorbol-13-acetate-promoted mouse skin papillomas are promotable by benzoyl peroxide.Mutat Res. 2004 Apr 14;548(1-2):35-45. doi: 10.1016/j.mrfmmm.2003.12.017. Mutat Res. 2004. PMID: 15063134
-
Mutually exclusive mutations of the Pten and ras pathways in skin tumor progression.Genes Dev. 2004 Aug 1;18(15):1800-5. doi: 10.1101/gad.1213804. Genes Dev. 2004. PMID: 15289454 Free PMC article.
-
Genetics of chemical carcinogenesis--II. Papilloma induction and malignant conversion in susceptible (Car-s) and resistant (Car-R) lines of mice produced by bidirectional selective breeding and in their (Car-S X Car-R) F1 hybrids.Carcinogenesis. 1994 Nov;15(11):2629-35. doi: 10.1093/carcin/15.11.2629. Carcinogenesis. 1994. PMID: 7955117
-
Non-melanoma skin cancer in mouse and man.Arch Toxicol. 2013 May;87(5):783-98. doi: 10.1007/s00204-012-0998-9. Epub 2012 Dec 25. Arch Toxicol. 2013. PMID: 23266722 Review.
Cited by
-
Role of carbonic anhydrases in skin wound healing.Exp Mol Med. 2017 May 19;49(5):e334. doi: 10.1038/emm.2017.60. Exp Mol Med. 2017. PMID: 28524177 Free PMC article.
-
Prophylaxis of Diallyl Disulfide on Skin Carcinogenic Model via p21-dependent Nrf2 stabilization.Sci Rep. 2016 Oct 19;6:35676. doi: 10.1038/srep35676. Sci Rep. 2016. PMID: 27759091 Free PMC article.
-
R-Ras deficiency does not affect papain-induced IgE production in mice.Immun Inflamm Dis. 2017 Sep;5(3):280-288. doi: 10.1002/iid3.168. Epub 2017 May 11. Immun Inflamm Dis. 2017. PMID: 28497586 Free PMC article.
-
Pathological Angiogenesis Requires Syndecan-4 for Efficient VEGFA-Induced VE-Cadherin Internalization.Arterioscler Thromb Vasc Biol. 2021 Apr;41(4):1374-1389. doi: 10.1161/ATVBAHA.121.315941. Epub 2021 Feb 18. Arterioscler Thromb Vasc Biol. 2021. PMID: 33596666 Free PMC article.
-
T-cell-expressed proprotein convertase FURIN inhibits DMBA/TPA-induced skin cancer development.Oncoimmunology. 2016 Oct 14;5(12):e1245266. doi: 10.1080/2162402X.2016.1245266. eCollection 2016. Oncoimmunology. 2016. PMID: 28123881 Free PMC article.
References
-
- Lowe D. G. et al. Structure of the human and murine R-ras genes, novel genes closely related to ras proto-oncogenes. Cell 48, 137–146 (1987). - PubMed
-
- Song J., Zheng B., Bu X., Fei Y. & Shi S. Negative association of R-Ras activation and breast cancer development. Oncol. Rep. 31, 2776–2784 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
