

Protein structural dynamics revealed by time-resolved X-ray solution scattering
- PMID: 26134248
- PMCID: PMC4659713
- DOI: 10.1021/acs.accounts.5b00198
Protein structural dynamics revealed by time-resolved X-ray solution scattering
Abstract
One of the most important questions in biological science is how a protein functions. When a protein performs its function, it undergoes regulated structural transitions. In this regard, to better understand the underlying principle of a protein function, it is desirable to monitor the dynamic evolution of the protein structure in real time. To probe fast and subtle motions of a protein in physiological conditions demands an experimental tool that is not only equipped with superb spatiotemporal resolution but also applicable to samples in solution phase. Time-resolved X-ray solution scattering (TRXSS), discussed in this Account, fits all of those requirements needed for probing the movements of proteins in aqueous solution. The technique utilizes a pump-probe scheme employing an optical pump pulse to initiate photoreactions of proteins and an X-ray probe pulse to monitor ensuing structural changes. The technical advances in ultrafast lasers and X-ray sources allow us to achieve superb temporal resolution down to femtoseconds. Because X-rays scatter off all atomic pairs in a protein, an X-ray scattering pattern provides information on the global structure of the protein with subangstrom spatial resolution. Importantly, TRXSS is readily applicable to aqueous solution samples of proteins with the aid of theoretical models and therefore is well suited for investigating structural dynamics of protein transitions in physiological conditions. In this Account, we demonstrate that TRXSS can be used to probe real-time structural dynamics of proteins in solution ranging from subtle helix movement to global conformational change. Specifically, we discuss the photoreactions of photoactive yellow protein (PYP) and homodimeric hemoglobin (HbI). For PYP, we revealed the kinetics of structural transitions among four transient intermediates comprising a photocycle and, by applying structural analysis based on ab initio shape reconstruction, showed that the signaling of PYP involves the protrusion of the N-terminus with significant increase of the overall protein size. For HbI, we elucidated the dynamics of complex allosteric transitions among transient intermediates. In particular, by applying structural refinement analysis based on rigid-body modeling, we found that the allosteric transition of HbI accompanies the rotation of quaternary structure and the contraction between two heme domains. By making use of the experimental and analysis methods presented in this Account, we envision that the TRXSS can be used to probe the structural dynamics of various proteins, allowing us to decipher the working mechanisms of their functions. Furthermore, when combined with femtosecond X-ray pulses generated from X-ray free electron lasers, TRXSS will gain access to ultrafast protein dynamics on sub-picosecond time scales.
Figures
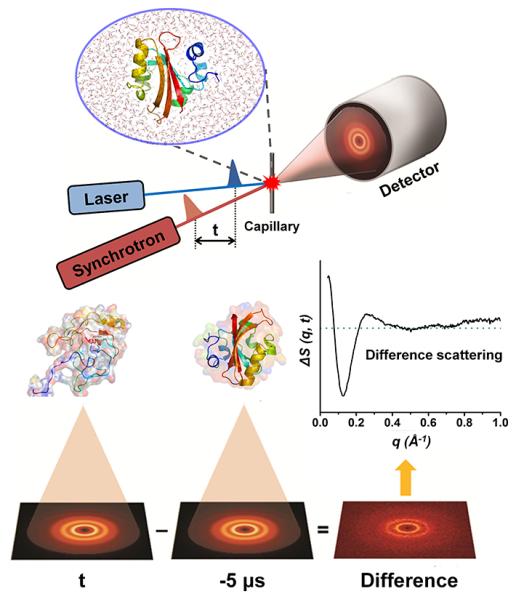



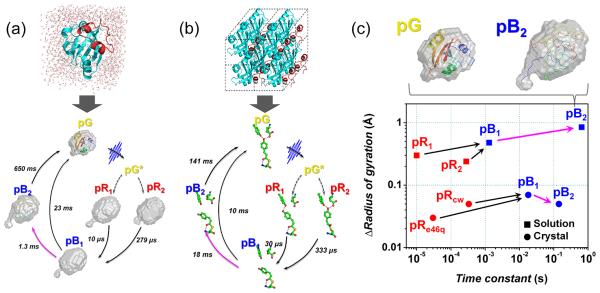


References
-
- Kukura P, McCamant DW, Mathies RA. Femtosecond stimulated Raman spectroscopy. Annu. Rev. Phys. Chem. 2007;58:461–488. - PubMed
-
- Kennis JTM, Larsen DS, Ohta K, Facciotti MT, Glaeser RM, Fleming GR. Ultrafast protein dynamics of bacteriorhodopsin probed by photon echo and transient absorption spectroscopy. J. Phys. Chem. B. 2002;106:6067–6080.
-
- Zanni MT, Hochstrasser RM. Two-dimensional infrared spectroscopy: a promising new method for the time resolution of structures. Curr. Opin. Struc. Biol. 2001;11:516–522. - PubMed
-
- Lewis JW, Goldbeck RA, Kliger DS, Xie XL, Dunn RC, Simon JD. Time-resolved circular-dichroism spectroscopy - experiment, theory, and applications to biological-systems. J. Phys. Chem. 1992;96:5243–5254.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
