

Genome Wide Binding Site Analysis Reveals Transcriptional Coactivation of Cytokinin-Responsive Genes by DELLA Proteins
- PMID: 26134422
- PMCID: PMC4489807
- DOI: 10.1371/journal.pgen.1005337
Genome Wide Binding Site Analysis Reveals Transcriptional Coactivation of Cytokinin-Responsive Genes by DELLA Proteins
Abstract
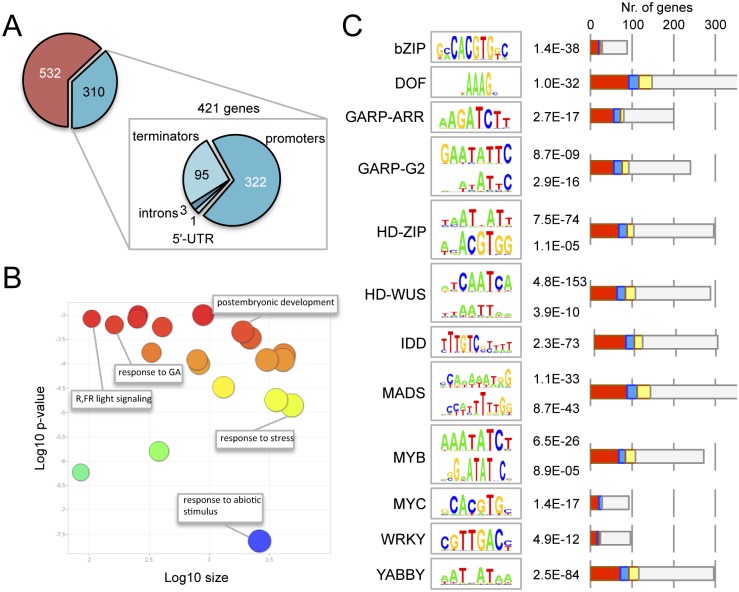
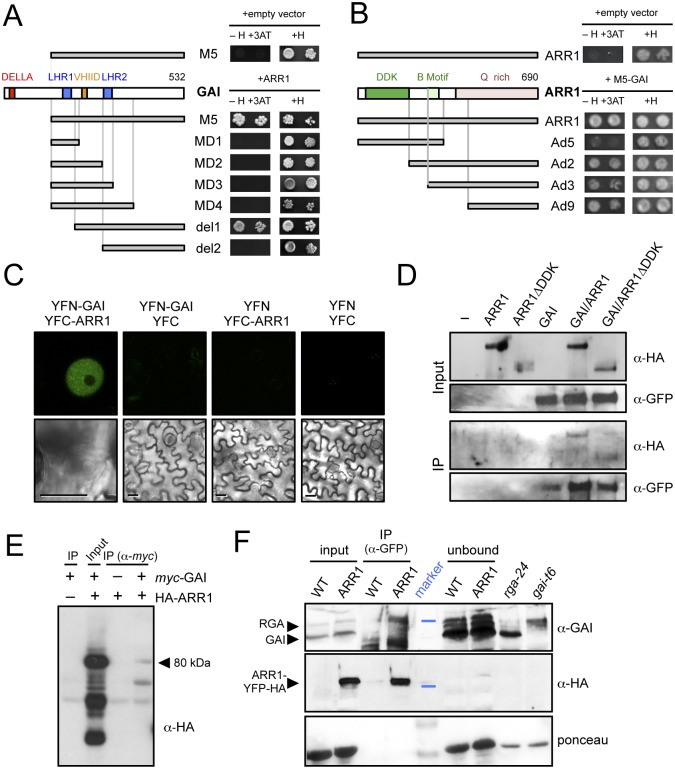
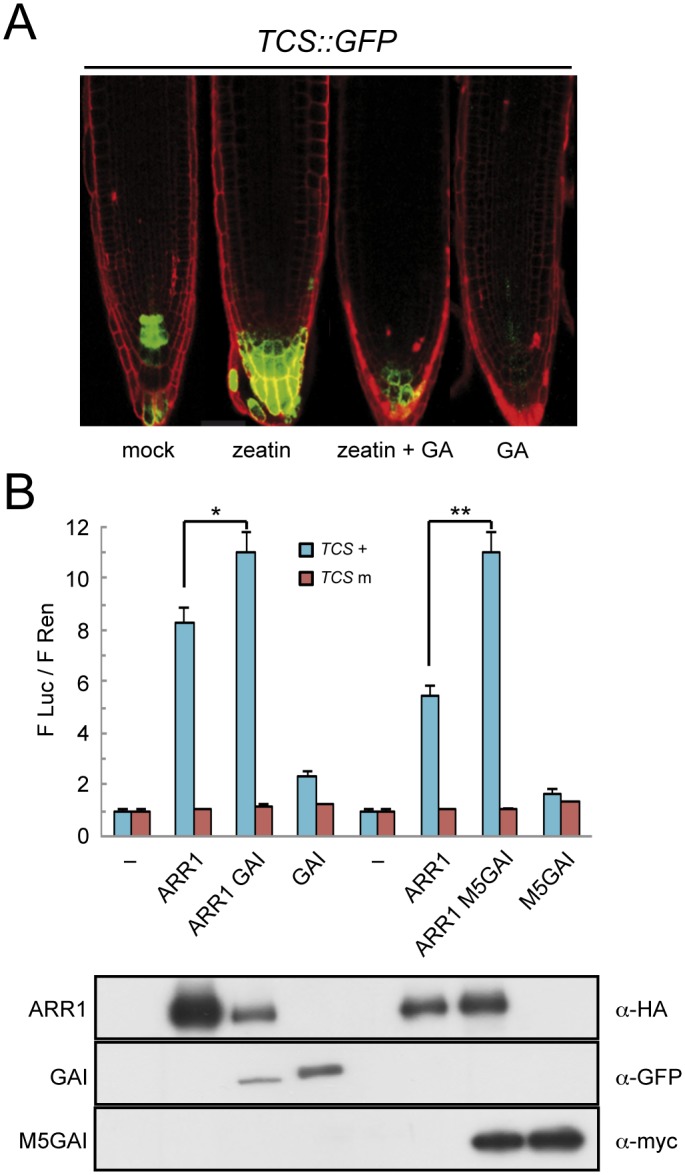
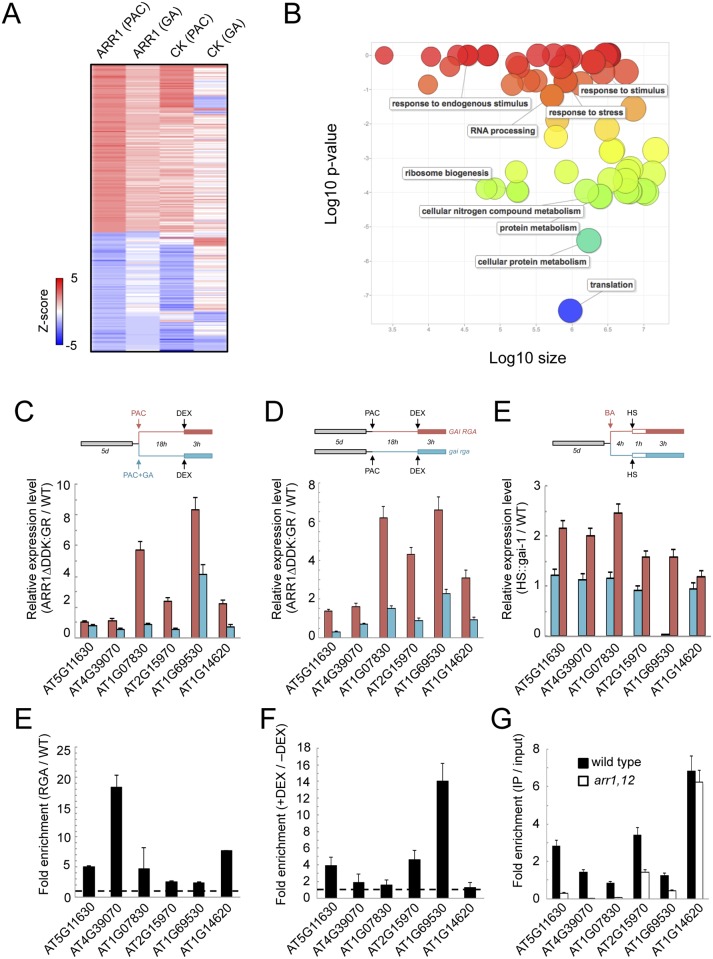
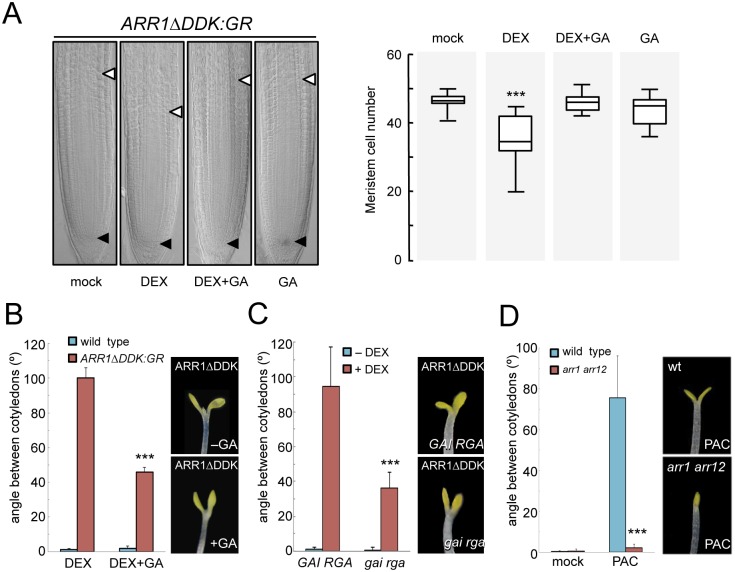
The ability of plants to provide a plastic response to environmental cues relies on the connectivity between signaling pathways. DELLA proteins act as hubs that relay environmental information to the multiple transcriptional circuits that control growth and development through physical interaction with transcription factors from different families. We have analyzed the presence of one DELLA protein at the Arabidopsis genome by chromatin immunoprecipitation coupled to large-scale sequencing and we find that it binds at the promoters of multiple genes. Enrichment analysis shows a strong preference for cis elements recognized by specific transcription factor families. In particular, we demonstrate that DELLA proteins are recruited by type-B ARABIDOPSIS RESPONSE REGULATORS (ARR) to the promoters of cytokinin-regulated genes, where they act as transcriptional co-activators. The biological relevance of this mechanism is underpinned by the necessity of simultaneous presence of DELLAs and ARRs to restrict root meristem growth and to promote photomorphogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The transcriptional repressor ARR1-SRDX suppresses pleiotropic cytokinin activities in Arabidopsis.Plant Physiol. 2008 Jul;147(3):1380-95. doi: 10.1104/pp.107.115436. Epub 2008 May 23. Plant Physiol. 2008. PMID: 18502977 Free PMC article.
-
Cytokinin induces genome-wide binding of the type-B response regulator ARR10 to regulate growth and development in Arabidopsis.Proc Natl Acad Sci U S A. 2017 Jul 18;114(29):E5995-E6004. doi: 10.1073/pnas.1620749114. Epub 2017 Jul 3. Proc Natl Acad Sci U S A. 2017. PMID: 28673986 Free PMC article.
-
In planta analysis of a cis-regulatory cytokinin response motif in Arabidopsis and identification of a novel enhancer sequence.Plant Cell Physiol. 2013 Jul;54(7):1079-92. doi: 10.1093/pcp/pct060. Epub 2013 Apr 24. Plant Cell Physiol. 2013. PMID: 23620480
-
Cytokinin signaling and transcriptional networks.Curr Opin Plant Biol. 2010 Oct;13(5):533-9. doi: 10.1016/j.pbi.2010.08.006. Epub 2010 Sep 17. Curr Opin Plant Biol. 2010. PMID: 20851038 Review.
-
Arabidopsis cytokinin signaling pathway.Sci STKE. 2007 Oct 9;2007(407):cm5. doi: 10.1126/stke.4072007cm5. Sci STKE. 2007. PMID: 17925576 Review.
Cited by
-
The pillars of land plants: new insights into stem development.Curr Opin Plant Biol. 2018 Oct;45(Pt A):11-17. doi: 10.1016/j.pbi.2018.04.016. Epub 2018 May 12. Curr Opin Plant Biol. 2018. PMID: 29763857 Free PMC article. Review.
-
Type-B response regulator OsRR22 forms a transcriptional activation complex with OsSLR1 to modulate OsHKT2;1 expression in rice.Sci China Life Sci. 2023 Dec;66(12):2922-2934. doi: 10.1007/s11427-023-2464-2. Epub 2023 Oct 30. Sci China Life Sci. 2023. PMID: 37924467
-
Meeting at the DNA: Specifying Cytokinin Responses through Transcription Factor Complex Formation.Plants (Basel). 2021 Jul 16;10(7):1458. doi: 10.3390/plants10071458. Plants (Basel). 2021. PMID: 34371661 Free PMC article. Review.
-
The ERF11 Transcription Factor Promotes Internode Elongation by Activating Gibberellin Biosynthesis and Signaling.Plant Physiol. 2016 Aug;171(4):2760-70. doi: 10.1104/pp.16.00154. Epub 2016 Jun 2. Plant Physiol. 2016. PMID: 27255484 Free PMC article.
-
DELLA-GAF1 Complex Is a Main Component in Gibberellin Feedback Regulation of GA20 Oxidase 2.Plant Physiol. 2017 Nov;175(3):1395-1406. doi: 10.1104/pp.17.00282. Epub 2017 Sep 15. Plant Physiol. 2017. PMID: 28916594 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/D522603/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E006922/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G023972/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E022618/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
